当代生物哲学研讨会(一) 自然选择主要作用于基因上吗?
Contemporary Philosophy of Biology
当代生物哲学研讨会
第一期
一场将近六十年的争论
自然选择主要作用于基因上吗?
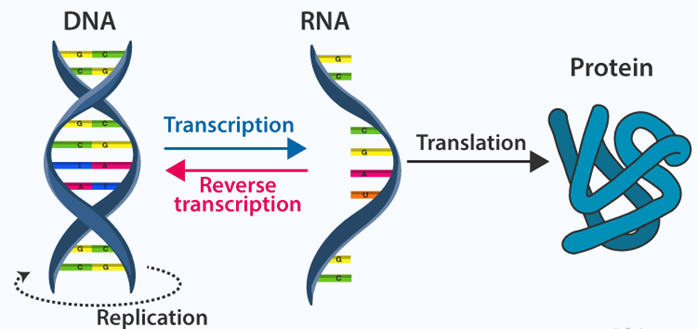
封面 DNA中心法则
图源自 https://byjus.com/biology/central-dogma-inheritance-mechanism/
在生命演化研究中,自然选择总是被反复提及和讨论。达尔文作为“自然选择”概念的提出者,对其做出了较为严谨的定义:自然选择是个体因表型差异而产生的生存和繁殖差异。它是生命系统演化的一个重要驱动力,即一个种群世代相传的可遗传特征的变化规则[1]。随着沃森和克里克关于DNA双螺旋结构的研究[2]发现,基因作为生物性状遗传的主要物质载体逐渐登上了生命科学的舞台。由此,关于自然选择是否主要作用于基因之上的争论便由此拉开序幕。
1966年,George C. Williams在《Adaptation and Natural Selection》[3](《适应和自然选择》)一书中,第一次正式提出了基因是选择的主要单元这一观点,即Gene’s-eye view的雏形。它强调个体基因传递在生物演化和行为中的重要性,并淡化了有机体本身受到的自然选择对演化的影响。

图1 George C. Williams (左)及其代表作Adaptation and Natural Selection封面(右)
在此之后,George的支持者Richard Dawkins为Gene’s-eye view这一观点提供了更多的论据。Richard认为基因是寄存于生物体的复制子,通过复制传递其基本模式和结构。该说法对于学过生物遗传学知识的高中生来说听起来很合理,然而争议的焦点在于:
虽然自然选择似乎是在个体层面上发挥作用,但真正有力的选择是否主要发生在基因层面上?
对于这个问题,支持者和反对者多年来开展了大量辩论。如1982年Elliott Sober 和Richard Lewontin两人在论文Artifact, Cause, and Genic Selection[4](人为因素、原因和基因选择)中就提出了对基因选择主义的反驳:
1. 除了基因层面,自然选择还可以作用于其他层次,包括个体和群体。例如,在群体选择理论中,群体的特征(如合作行为)可能对其成员的生存和繁衍产生积极影响。在3%到8%的鸟类中(包括家雀、普通秧鸡、橡果啄木鸟和灰驼鹨),它们在自己还没有繁殖的情况下会帮助父母抚养下一窝后代;生活在洞穴里的老鼠遇到天敌后不是自己先逃跑,而是发出叫声吸引天敌,警告群众。这些行为尽管会导致个体自身生殖资本下降,于自身不利,但是却可以提升群体整体适合度 ,说明自然选择除了基因,还可以作用于群体。
2. 基因选择主义忽视了表型的重要性:基因选择主义者过分强调基因在演化过程中的作用,而忽略了表型特征在自然选择过程中的重要性。基因和表型之间存在复杂的相互作用,仅仅从基因选择的角度很难完全解释生物体的形态和行为。
3. 基因选择主义未能充分考虑遗传因素与非遗传因素之间的相互作用。例如,在表型可塑性中,环境因素可能影响个体的表型,但并不会导致基因的改变 。也就是说基因选择主义可能无法解释这些表型变化背后真正的生物学机制。

图2 Elliott Sober(左) 、Richard Lewontin(中)及其论文Artifact, Cause, and Genic Selection封面(右)
没过多久,基因选择主义的支持者们也拉起了反攻的号角。1988年,Kim Sterelny和Philip Kitcher在论文The Return of the Gene[5](基因的回归)中回复了Sober和Lewontin的论点:
1. Gene’s-eye view可能不是描述演化过程的最佳理论,但并不意味着基因视角完全没有价值。早在上世纪80年代,许多与特定表型特征(如遗传性疾病)相关的基因就已经成功被鉴定,而且其中基因如何影响生物体的表型的遗传基础也逐渐被揭示。通过对这些特定基因进行编辑,人们还可以观察到生物体表型的变化,说明基因确实在生物体内扮演着关键角色。
2. Sober和Lewontin的观点将基因视角与表型选择和群体选择之间看作是一个二元对立的关系,然而在许多情况下,基因视角和其他选择层次之间的关系可能是互补的,而非竞争。虽然Sterelny和Kitcher承认Sober和Lewontin对基因视角的质疑在遗传与非遗传因素互作层面是合理的,但他们依旧认为基因视角在演化生物学中仍具有重要价值,特别是在遗传学和分子生物学的新发现、选择模型的改进以及演化发育生物学的发展等方面。这些因素为基因视角提供了新的论据和证据,从而为其在演化生物学中的地位辩护。
就这样,生物学家们关于Gene’s-eye view的论证持续了接近半个世纪。晦涩的论点与乏味的论据逐渐充斥了整场辩论。直到2006年,Samir Okasha为这场争论引入了哲学思辨的元素。在论文The Levels of Selection Debate: Philosophical Issues[6]中,他抛出了三个问题,将整场争论推入了高潮:
如何确定基因对生物体表型的因果贡献?
选择作用可能同时作用于多个层次,基因一定是主要单位吗?
自然选择和遗传漂变如何共同作用以塑造演化过程?
这是三个难以用三言两语回答的问题,Okasha提出这些问题时也想到了短时间内很难得到解决与解释,于是他将目光拉长,期待在未来得到自己问题的答案。
在2023年的今天,我们总结了近十几年科研工作者们关于自然选择是否主要作用于基因层面上观点的相关研究进展,尝试将这场辩论正反两方观点的研究现况进行梳理。
图3 争论现况
正方:
自然选择能直接作用于基因/DNA序列层面
当我们把关注点放到自然选择是否作用于单个基因层面这一问题时,事情开始变得有趣起来。以医院中对各种抗生素具有耐药性的微生物为例:抗生素的选择压力直接作用于编码抵抗抗生素的基因产物,通过杀死那些不能产生抵抗抗生素物质的细菌,将具有耐药性的微生物选择下来,这便是自然选择作用于单个基因的经典案例。当然我们知道调控性状背后往往不止一个基因在发挥作用,而且这些不同基因的作用效果却有区别。例如一个种群的基因组中存在20个变异基因,可能其中两个基因就贡献了50%效应,而剩下的18个基因贡献了剩下的50%,在这种情况下,选择可能会对这两个基因产生强烈的影响,而对其余的基因则产生微弱的影响,或者根本不产生影响。从全基因组关联分析角度来说,如果这些性状真的是数百个基因共同作用的结果,就很难找到对这些表型有显著影响的基因。另外,如果同时控制某个优势性状的位于不同染色体的三个基因,需要将该优势保留的话,配子中同时保留这三个基因的概率是1/2 × 1/2 × 1/2 = 1/8,而如果是连在一起(连锁),概率就是1/2,这比1/8的概率大的多,也容易发生的多。因此,正方观点认为自然选择作用的往往是单个或连锁的几个基因,而不是独立的多个基因。
除了上述提到的正方观点中“自然选择能直接作用于基因”,Carmen Sapienza将作用对象进一步扩展到DNA序列中,Carmen Sapienza认为:今天的大部分基因组结构是自然选择直接作用于DNA序列的结果[7]。也就是说,他认为在自然选择的压力下,DNA序列片段会在基因组内如同寄生虫一般自我复制,使得自身在细胞分裂/减数分裂之前有足够多副本,从而保证序列的延续。因为我们要知道,对于生物表型没有任何贡献的“垃圾序列”,它们在人类基因组的数量也已经超过了三分之一,尽管它们并不受表型压力,却依然被选择了下来。
以人类中ALU元件为例,在极少数情况下,一个元件插入到基因附近可能会影响该个体表型,但实际上人类中却存在上百万个ALU元件,而且它们中的大多数并不存在于基因内或基因附近。这说明在没有任何对生物体表型起作用的选择压力之下,这些转座元件绝大多数却能够在基因组中得以选择和保留,甚至大量增加。
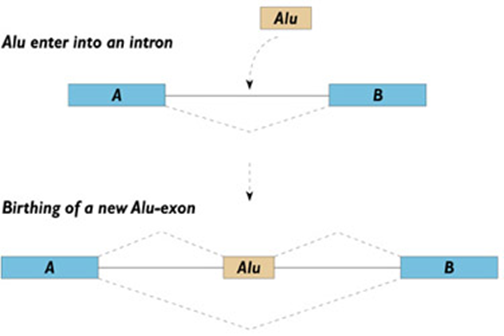
图4 转座子插入基因图示
图源自https://hoopermuseum.earthsci.carleton.ca/neanderthal/n-alu.html
对于自然选择直接在DNA序列水平上起作用的这一论点,最令人信服的证据也许是减数分裂驱动(meiotic driver)的例子,即染色体在减数分裂时的非随机分离。我们知道雌性并不是产生四个大小和遗传含量相等的配子,而是产生一个大配子(卵子)和两个较小的减数分裂产物(第一和第二极体)。Wu and Sapienza[8]发现单个染色体的变异可能会影响其分离到卵子的概率,从而提升其传递到下一代的可能性。换句话说,减数分裂过程中,着丝粒通过与纺锤体相连,将染色体迁移到指定极点,但并不是所有着丝粒都能与纺锤体相连,从而使得染色体被“拉向”卵细胞一侧。研究表明,当着丝粒区域有更多的串联重复DNA序列时,染色体更容易被纺锤丝迁移到卵子中[9]。
在染色体层面上,自然选择除了作用于着丝粒,还可能通过罗伯逊易位实现。罗伯逊易位是发生在近端着丝粒染色体的一种易位形式,当两个近端着丝粒染色体在着丝粒部位或附近部位发生断裂后,二者的长臂在着丝粒处结合在一起,形成一条由长臂构成的衍生着丝粒;两个断臂则构成一个小染色体,它往往在第二次减数分裂时丢失(图5)。罗伯逊易位并不直接作用于表型,但是这种变异已被证明是哺乳动物演化中生殖隔离的主要机制。这些染色体变异的产生频率非常高——在人类女性减数分裂中约为千分之一。如此高的变异率如果对人类演化没有优势,这些染色体变异为什么还会出现和被选择?
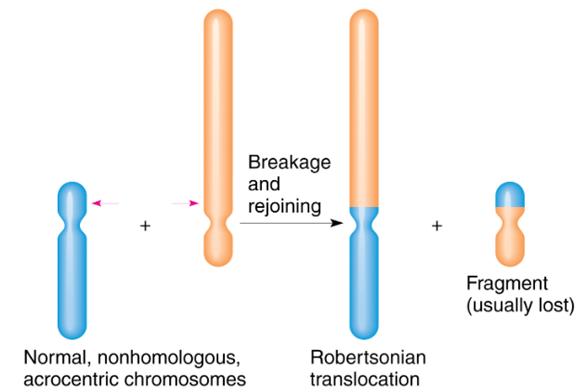
图5 罗伯逊易位示意图[10]
可见这些直接对DNA水平进行的选择效应影响着物种的生殖适应性,而生殖适应性往往是任何群体遗传学讨论中最重要的结果衡量标准。当我们评判对某个性状的选择优势程度时,就需要用适应度来衡量,即产生后代的数目。通过对冰岛人数据统计发现,适应度与重组率有关,即有更高重组率的妇女能够在更大的年纪生育,且有更多的后代,而影响重组率的往往是单个位点、单个基因或少数基因[11]。
图6 重组的发生(左)与高生育力家庭(右)
左图源 https://isogg.org/wiki/Recombination,
右图源自谷歌
反方:
自然选择并不主要作用于基因层面
有人支持,也自然有人反对,生物学上关于很多问题的争论从来都不是非黑即白的针锋相对,而是相辅相成的互利影响。基因选择学说的反对者Richard M. Burian就旗帜鲜明地提出了自己的质疑,他认为基因选择学说太过武断,如果我们将视角聚焦于自然选择的因果过程,就会发现并不是所有的遗传都是通过基因来实现的。William Wimsatt于1980年对此进行了论述:
当两种不同基因同时发生变异时,选择的发生依赖于基因频率的变化,即其中一个等位基因是否被选择取决于两个位点上等位基因在群体中的比例。在这种情况下,选择是否有利于或不利于一个特定的等位基因可能取决于出现在另一个位点的等位基因的频率,其结果是一个动态过程,由性状与生态环境关联,以及两个基因及其产物或效应的因果关系驱动,但总的来说,选择并不偏爱任何一个基因变体。由于季节性和零星的变化可能会极大地影响另一个位点上等位基因的比例,因此在种群具有特定组成的情况下,无法从特定等位基因的选择优势预测演化结果。
以上论述其实指出了关键点:如果对基因型-表型关系以及相关的选择制度没有清晰和正确的解释,人们就不能武断确定频率增加的单个等位基因是否是选择的主要目标。即使承认性状有很强的遗传因果关系,也不能由此认为对一个基因及其性状的分析足以解释所讨论生物的演化轨迹。
以镰状细胞性贫血为例,在接触携带疟疾寄生虫蚊子的人群中,血红蛋白基因的镰状细胞等位基因很普遍。在没有医疗干预的情况下,携带该等位基因纯合子的个体几乎总是在进入青春期之前因镰状细胞性贫血而死亡。尽管如此,该等位基因在疟疾地区保持在相当高的频率,因为它为人类杂合子提供了相当大的抗疟疾能力,镰状细胞运输的氧气可以足以低到让疟原虫无法生存,因此尽管镰状细胞性贫血死亡代价巨大,抵抗疟疾带来的净效果依然更有利(有点损敌一千自损八百的意思)。
在这种背景下,Elliott Sober和Richard Lewontin也于1982年提出了另一个反对基因选择学说的观点:自然选择并不主要作用于等位基因序列的改变,而是根据不同疟疾程度的环境与对青少年的死亡威胁程度之间的动态平衡关系来决定不同血红蛋白在血细胞中的分布的效应,即该等位基因在群体中的频率(而在抗生素的例子中,有无抗生素的环境是直接作用于编码抵抗抗生素蛋白的基因,去选择基因的有无;但是在镰状细胞性贫血中,有无疟疾的环境并不是直接作用于编码血红蛋白的基因,而是根据血红蛋白的含量分布来决定该等位基因的频率分布,而该过程是动态的,并不只针对个体本身,更侧重于对群体的选择)。
总体而言,该论点认为随着时间的推移,选择作用于种群,增加了种群中因果相关的基因型的频率。也就是首先需要弄清楚是基因还是基因型如何与表型相关。为了弄清楚因果关系,我们必须进一步探究种群中可用的基因型以及基因型和表型随时间变化的动态。
在蚜虫中也存在着非常有趣的现象,即过冬时的雌性(AA+XX)蚜虫与雄性(AA+XO)蚜虫在上个夏季末交配,然后的卵于来年春天孵化,其后代均为雌性(AA+XX)。这些雌性会通过孤雌生殖产生一系列全雌性的后代,通常会存活三到四周,这些后代并不产卵,但是会产生已经是胚胎的后代。尽管这些雌性和它们的雌性后代在基因上相同,但它们的饮食差异却可以改变它们下一代及下下一代的大小、形态和生育能力。例如,它们可以确定这些后代是否有翅膀(寻找另一个寄主植物)或无翅(留在同一个寄主上)。夏末在光线、营养和温度信号的刺激下,这些雌性就会生出有翅膀的雄性和有性的雌性,他们于夏末交配,产的卵又进入下一个繁殖周期 (图7)。
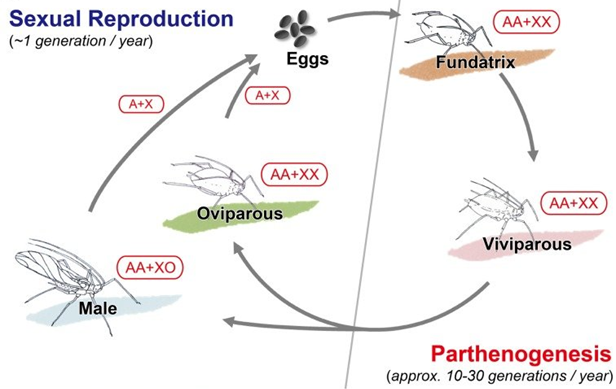
图7 蚜虫生活史[12]
从刚刚所描述的蚜虫的生活史中,我们可以得到两个论点:(A)有机体的后代的特征是由环境和遗传因素共同决定的。某些看似遗传的特征,例如有性生殖的雌性的生育能力,或者一个孤雌生殖的雌性的女儿有翅膀的比例,可能不是由它的基因决定的,而是由它的营养状况和它当季的母系血统决定的。(B)选择是偶发性的,它不仅仅作用于基因,还可以直接作用于表型,但它如何起作用必须在适当尺度的整个周期中衡量。
因此,反方认为选择通常不是作为一种单独的力量作用于一个孤立的基因或特征,而是作为一种平衡装置,在长时间内以分散的事件对多个特征产生影响。维持多个性状的选择压平衡是常态,而不是单一的力量影响一个孤立的基因(或性状)。
结语
选择确实可以直接作用于基因、表型和群体,但是哪个才是主要的作用单元、哪一种选择更普遍是存在争议的。站在正方的角度,基因是那个因,是几乎所有遗传形式的基础,而表型只是果,从基因到表型的顺式调控关系是非常清晰的。打个比方就是,选择的这个过程就像复杂的化学反应,但是其中在大多数过程中都有“速率限制”步骤,尽管每一步都很重要,但是基因突变比其他步骤的突变具有更大的表型效应。站在反方的角度,有机体的表型和表达的蛋白不仅仅是果,它也可以主动的去改变环境,从而改变调控网络,在不涉及DNA序列的情况下遗传。但是这种现象是普遍存在的吗?这种逆向改变、可遗传的机制是尚不明了的,其遗传的载体到底是什么?也许当我们能够回答这些问题的时候,该争论才会有答案。
参考文献
[1] Darwin C. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life[J]. London: Murray, 1859.
[2] Watson J D, Crick F H C. Molecular structure of nucleic acids: a structure for deoxyribose nucleic acid[J]. Nature, 1953, 171(4356): 737-738.
[3] Williams G C, Burt A. Adaptation and natural selection[M]. na, 1966.
[4] Sober E, Lewontin R C. Artifact, cause and genic selection[J]. Philosophy of science, 1982, 49(2): 157-180.
[5] Sterelny K, Kitcher P. The return of the gene[J]. The Journal of Philosophy, 1988, 85(7): 339-361.
[6] Okasha S. The levels of selection debate: philosophical issues[J]. Philosophy Compass, 2006, 1(1): 74-85.
[7] Dawkins, R. The selfish gene [M]. New York: Galaxy Books. 1976.
[8] Wu, G et al. Maternal transmission ratio distortion at the mouse Om locus results from meiotic drive at the second meiotic division [J]. Genetics, 2005, 170, 327–334.
[9] Nikalayevich E, Verlhac MH. Selfish centromeres, selfless heterochromatin [J]. Cell. 2021, 184, 4843-4844.
[10] PJ Russell. iGenetics: A Molecular Approach (3rd Edition), 2010. Benjamin Cummings, San Francisco.
[11] Kong, A. et al. Recombination rate and reproductive success in humans [J]. Nature Genetics, 2004, 36, 1203–1206,.
[12] Ogawa K, Miura T. Aphid polyphenisms: trans-generational developmental regulation through viviparity. Front Physiol. 2014, 5:1.








