SA论文解读:跨物种杂交驱动猕猴新物种起源
从达尔文时代开始,新物种的形成方式一直都是演化生物学最关心的话题之一。来自中科院昆明动物所等多个研究单位的团队对12种猕猴的基因组进行了系统发生和比较基因组学分析研究,发现跨物种杂交是驱动猕猴新物种形成的重要因素。该研究是灵长类基因组计划的阶段性成果之一,近日已在知名学术期刊《科学进展》(Science Advances)上以题为“Comparative genomics reveals the hybrid origin of a macaque”(比较基因组学揭示了一类猕猴的杂交起源)的进行发表。
物种间杂交可能促进新物种的形成
在经典演化理论中,某个祖先种群因为迁徙和环境变化等因素被隔离成了两个无法互相交流的独立种群,它们各自慢慢累积了不同的突变,直到它们即便再次相遇也无法繁衍出可育后代,这就标志着新物种的形成。因此,达尔文把演化的历程描述为共同的祖先种群随时间不断推移不断分出不同枝杈的树状结构,也就是演化树。这种每个分支互不交叉的树状结构向我们表明每个现生物种的任何历史阶段的祖先都是来自于同一个种群。
除了经典演化理论中由地理隔离导致的生殖隔离之外,后来,人们逐渐意识到跨物种杂交也是新物种形成的机制之一。如在植物演化过程中,杂交一直被认为是物种形成的一种重要模式。而涉及到动物演化历程时,人们曾以为杂交形成新物种过去很少在动物中发生。但随着越来越多物种的基因组数据被整合起来,人们发现那些经历了快速适应辐射的物种群体易于杂交,包括蝴蝶、犬类、狒狒、鸟类和熊,因为这些物种之间的不亲和性很弱。也就是说,在它们的演化历程中,某些祖先来自于多个不同种群的杂交,而非一直都是某个单一种群的后代。这时候,简单的树状结构实际上并不能准确概括真实的演化历程。
作为最成功的灵长类物种之一,猕猴属(Macaca)很是研究种间杂交和物种形成之间相互作用的一个很好的模型。猕猴现存23个物种,广泛分布在南亚、东亚和东南亚,唯一的例外是仅限于北非阿特拉斯山脉的西尔瓦纳斯猕猴(M. sylvanus)。此前的古生物学和分子数据表明,猕猴起源于大约700万年前的北非,然后在过去的500万年里在亚洲经历了物种形成的爆发。尽管属内某些不同种的猕猴在形态和行为上差别很大,但它们的地理分布存在广泛的连续性,这使得分布区域有重叠的不同猕猴物种之间都有潜在的杂交可能性。而某些猕猴物种如束尾猴和短尾猕猴表现出独特的混合表型,但杂交的更广泛的作用及其具体效果很大程度上仍然是未知的。于是,通过多基因组分析来确定猕猴属的物种演化历程,评估种间杂交在物种形成中的作用,并探索猕猴物种在混合表型上的遗传基础成为了一个很好的选择。
食蟹猴种组有情况
研究者们首次对分布在不同区域的10种猕猴进行了基因组测序和组装,并整合了两种过去已完成测序的猕猴,共使用12种猕猴的基因组进行系统发生和比较基因组学分析研究。
本研究分析的12种猕猴的地理分布情况。猕猴图片版权为国际自然保护联盟物种生存委员会灵长类专家组,©2013,Stephen D. Nash所有
为了推断出猕猴物种间的正确分支模式,并表征跨基因组的异质系统发育信号,研究者们将猕猴基因组划分成许多个50kb的DNA序列片段(这些片段被称为基因组窗口),然后对每个基因组窗口序列在所有猕猴之间的比对结果通过最大似然法,构建一棵窗口树。然后将许多窗口树整合成一个整体的物种演化树。虽然整体的物种树与过去对猕猴种间关系的基本认知一致,但研究人员发现,组成它的不同窗口树之间存在大量的拓扑结构是不一致的。
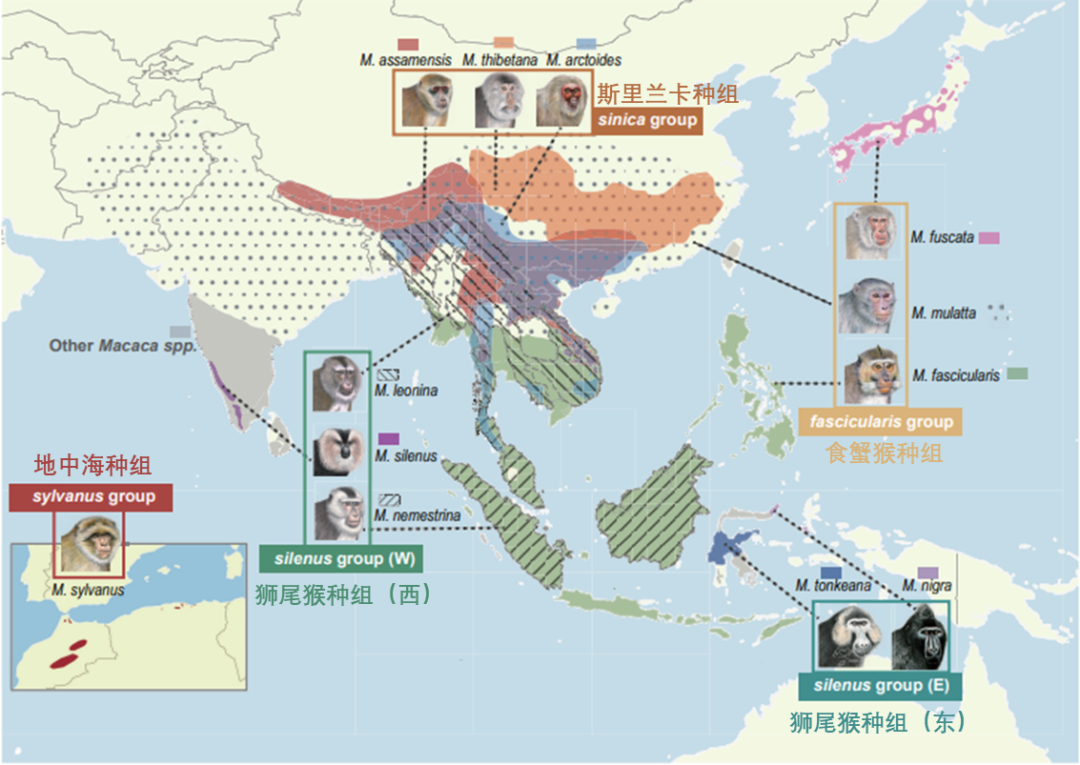
为了更好地理解这些系统发育的不一致在猕猴群体之间的相互关系,研究者们根据亲缘关系和特征将12个猕猴物种划分为5个种组进行研究。而在这个过程中,食蟹猴种组的位置出现了摇摆不定的情况,其中有两种拓扑结构占主导地位:一种支持食蟹猴种组(fascicularis group)是斯里兰卡种组(sinica group,主要分布在斯里兰卡等地)的姐妹种组(T1),而另一种(T2)支持食蟹猴种组是狮尾猴种组(silenus group)的姐妹种组。
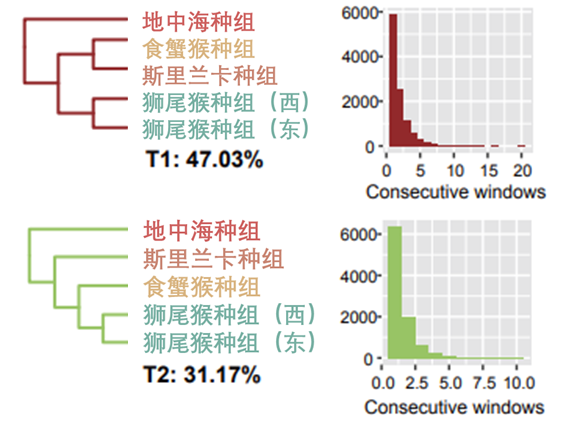
两种主要拓扑结构的分布情况(中科院昆明动物所吴东东课题组提供)
如此情况,研究人员推测食蟹猴种组可能起源于一个古老的杂交,它是由狮尾猴种组的祖先与斯里兰卡种组的祖先杂交而来的。
构建系统发生网络验证杂交起源假说
如果食蟹猴的杂交起源假说是正确的,我们就应该会看到杂交谱系和两个亲本谱系之间观察到同等水平的序列差异。为了验证这一点,研究者们分别比较了常染色体和X染色体上食蟹猴种组相对于狮尾猴种组和斯里兰卡种组的序列差异和相对差异时间。仅用性染色体数据计算的这两个统计量没有明显差异;而对于常染色体,食蟹猴与另两个种组之间的平均序列差异值(0.1616比0.1631)和平均估计年龄(3.45Mya比3.56Mya)只有轻微的差异。也就是说,X染色体和低重组区域在亲本谱系中表现出几乎相同的贡献。这表明食蟹猴种组受随后回交的影响较小,因此可能在保持杂种完整性方面发挥了重要作用。在排除了其他可能产生这种不一致特征的机制,如不完全谱系分流(ILS)和成种后的定向基因流动之后,研究者们确认观察到的模式与杂交成种情景下的预期模式一致。
为了进一步弄清关于其在初始杂交中的方向的信息,研究人员接下来对线粒体和Y染色体序列(长度约480kb)进行系统发育分析(前者代表母系遗传后者代表父系遗传),以确定杂交模式。他们在线粒体和Y染色体数据之间发现了明显的拓扑差异,其中食蟹猴种组的线粒体基因组类似于斯里兰卡种组,Y染色体类似于狮尾猴种组。这表明,古老的杂交主要或仅发生在原斯里兰卡种组的雌性和原始狮尾猴种组的雄性之间。
于是,在综合了12种猕猴的基因组数据的比较结之后,研究者们建立了一个揭示猕猴属的复杂物种起源关系的系统发生混合网络。这有别于传统演化理论中各枝互不交叉的简单树状结构,该演化网络向我们展示了物种间基因交流的丰富信息。
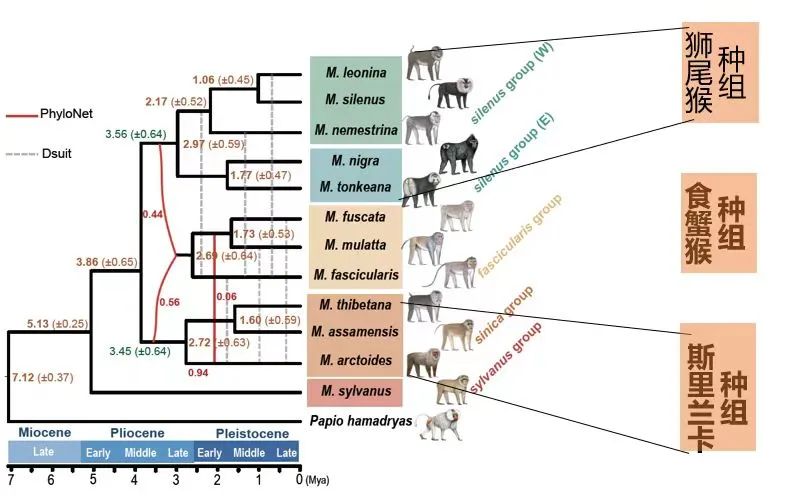
通过基因组比较建立的猕猴属内物种起源关系的系统发生网络,该网络揭示了食蟹猴的杂交起源(中科院昆明动物所吴东东课题组提供)
探索食蟹猴混合表型的遗传基础
评价杂交物种形成是否成功取决于这个新物种在生理形态上是否产生了独特的,稳定的遗传结构,并与其祖先物种形成生殖隔离。显然食蟹猴种组的几个独立物种是符合这一标准的。今天,由于生态地理障碍或行为差异,食蟹猴种组的各物种与它们的两个亲本种组分离得很好。然而,由于其起源于跨种杂交,现存的食蟹猴种组成员均显示出其亲本物种形态特征的独特混合模式。例如,食蟹猴的性器官和性皮肤整体上表现为其两种亲本的组合。这种混合的性状特征大多与生殖系统有关,因此研究者们猜测与这些特征相关的基因也应在假定的亲本谱系中表现出马赛克模式(即一部分更像“父系祖先”狮尾猴种组,另一部分更像“母系祖先”斯里兰卡种组)。
为了确定这些混合表型的遗传机制,他们使用了最新的分析方法来检测在食蟹猴和它们的“亲本物种”之间的正选择基因。食蟹猴基因组中,分别有216和195个正选择基因来自狮尾猴和斯里兰卡种组。前者中的7个基因携带斯里兰卡种组特有的非同义突变(“母系突变”),后者中则有4个基因携带狮尾猴种组特有的非同义突变(“父系突变”)。其中一个携带“母系突变”的正选择基因UGT1A9基因编码了一个促进雌激素降解的酶,研究者们推测这个在食蟹猴及其“母系种群”斯里兰卡猕猴之间的共同基因突变可能有助于通过影响雌激素水平来减少两组雌性的性皮肿胀。综上所述,这些结果提供了新的遗传证据来支持在食蟹猴种组中观察到的生殖形态的马赛克模型。
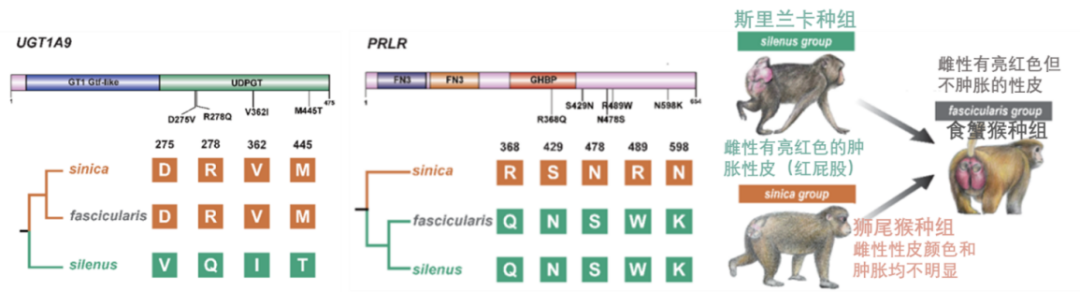
食蟹猴种组混合表型的遗传机制解释。左:携带母系突变的食蟹猴正选择基因UGT1A9,中:携带父系突变的食蟹猴正选择基因PRLR,右:食蟹猴的性皮肿胀表型处于斯里兰卡猕猴和狮尾猴的状态之间(中科院昆明动物所吴东东课题组提供)
猕猴属演化网络带来的更多新发现
研究者们构建了演化网络之后进一步发现了新的物种分支模式。如在狮尾猴种组内不同物种之间的关系上,人们曾因栖息地分布相近、所处生态环境较为类似,以及表型也很类似而认为南平顶猴(M. nemestrina)和北平顶猴(M. leonina)互为姐妹物种。然而根据这项研究所构建出来的更完善的系统发生关系网络来看,来自南印度西高止山脉的狮尾猕猴(M. silenus)和北平顶猴才是演化关系上更为接近的姐妹物种。过去我们发现,南平顶猴和北平顶猴是已知的东半球非人灵长类动物中唯二易感染HIV-1病毒的物种,这是因为其细胞中表达一种新型 TRIM5-CypA2 融合蛋白,这种突变使得TRIM5蛋白无法限制 HIV-1病毒的复制。这也是使人们曾对于狮尾猴种组系统发生模式出现错误理解的因素之一。根据同源序列分析,研究者们在狮尾猕猴中发现了相同的TRIM5-CypA2融合基因现象,这表明这种逆转录事件在至少217万年前西方狮尾猴种组(W)的共同祖先中就已出现。外周血单核细胞(PBMCs)体外感染实验证实,狮尾猕猴确实可以感染HIV-1病毒。
综上所述,在基因组时代之前,我们对于物种演化的理解往往是片面地来源于对少数特征的跨物种比较。而全基因组分析则能为我们带来更深刻、更全面的演化认知。这项研究向我们充分展示了物种间的演化关系以及新物种的形成过程可能远非单一树状结构所描述的那样简单,不断分叉又不断汇聚的演化轨迹交织出远比预想更复杂绚烂的演化网络图景。在此过程中复杂的基因交流关系也提供了除随机突变之外的更多表型多样性来源。相信会有越来越多曾基于片面信息而得出的物种演化认知将在未来被更全面的大数据组学研究所改写。








