铭刻在基因组上的演化史诗:从蛇为什么没有脚谈起
历史上,蛇类与我们人类的祖先打了不知多久的交道,以至于各大古文明大多都有与蛇类相关的传说,例如我们中华民族的精神图腾——“龙”,就是以长着爪子的大蛇为基础模板想象出来的。可我们都知道,蛇类是没有四肢的,两千多年前,我们中国人的老祖宗就留下了“画蛇添足”的典故。可能有许多人都曾好奇过,蛇类为什么长不出四肢?这个问题,也许能通过evo-devo的研究角度给出答案。
01. 什么是evo-devo?
Evo-devo是演化发育生物学(evolutionary developmental biology)的缩写。传统的进化学家们主要关注基因突变为生物在特定环境下的生存繁衍带来什么样的影响,更强调外因对演化的影响(例如蛇类失去四肢有什么好处),而evo-devo关注基因组变异通过影响哪些胚胎发育过程来改变物种的形态和生理结构,(例如蛇类基因组的哪些突变破坏了四肢发育过程)更强调内因对演化的影响。2016年,Axel Visel团队的研究在基因组层面上解答了这个问题,简单地讲:仅仅十几个碱基的突变就直接决定了蛇类四肢发育的失败,此项成果发表在《Cell》杂志上[1]。
四肢发育的成功取决于一种叫音猬因子(Sonic hedgehog, SHH)的形态发生素(morphogen),所谓形态发生素,就是一类在胚胎发育的特定时间特定位置由特定细胞分泌出来,用以诱导周围细胞发生分化的信号分子蛋白。这像极了你老板发给你催你加班的邮件。正如一家大公司正常运行离不开频繁的信息交流来确定每个基层员工的任务,胚胎发育过程中也是不断需要有各种各样的信号分子来规定每个细胞该朝什么方向分化或运动。多细胞生物的生理复杂性源自其发育过程中分化出来的复杂多样的细胞种类,而细胞分化则是不同基因在胚胎不同阶段不同区域差异表达的产物,因此对于基因时空特异性表达的调控问题的研究是evo-devo科学家们的唯一关切。那Evo-devo科学家们又是怎么解释蛇类进化过程中的四肢缺失问题呢?
02. Evo-devo破解四肢发育核心代码
在脊椎动物中,Shh基因在肢芽(胚胎中四肢的原始雏形)形成阶段表达,促进了肢芽的形成和进一步发育。这是因为Shh基因上游远端(约85万碱基距离)有一个叫做ZRS增强子的DNA序列,增强子是一种不编码基因的DNA序列,可一旦它被特定的蛋白结合,就会起到驱动或强化附近基因表达的功能(你可以把它想象成为控制灯泡的电路开关)。ZRS增强子是一种四肢特异性增强子,它起到了只在肢芽特异性地激活Shh基因表达的功能。而Visel团队通过比较不同脊椎动物的ZRS增强子后发现,蟒、蚺等更加古老的基础蛇类的ZRS增强子与四肢蜥蜴的直系同源区域共享约 80%同源序列,因此它们至今仍保留着微弱的后肢发育能力;但眼镜蛇、蝰蛇等高等蛇类的ZRS增强子则有更多的差异序列,它们也早已彻底失去了四肢发育的能力(图1)。因此作者认为,“虽然几乎所有被检查的蛇种都有ZRS增强子,但这种增强子进化约束的丧失与从基础蛇向高级蛇过渡时肢体结构的完全丧失相吻合”。
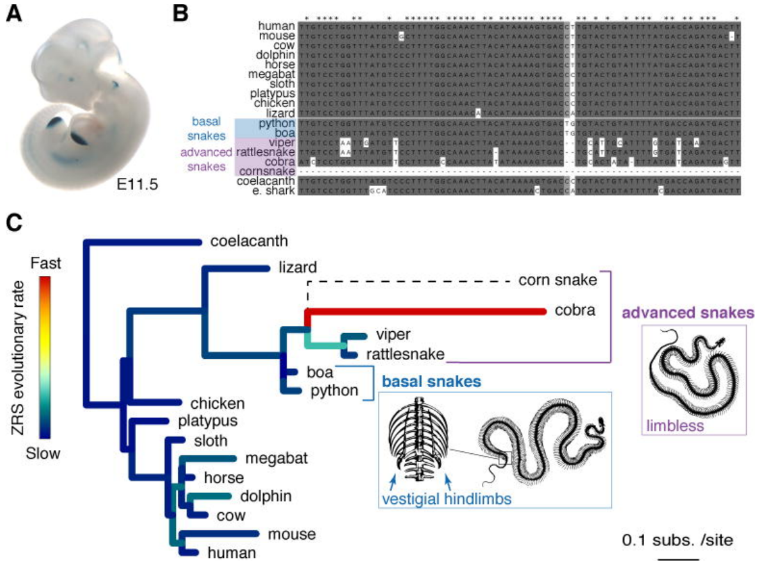
图1 脊椎动物树上肢体增强子的进化
为了验证自己的结论,Visel等人将不同物种的ZRS增强子替换下小鼠自身的ZRS,然后发现将蟒蛇或眼镜蛇的ZRS增强子转基因小鼠与ZRS敲除鼠的性状表现一致,即四肢发育严重受阻,而其他物种ZRS增强子替代小鼠则无明显异常(图2)。这说明了蛇类的四肢发育能力缺失的确源自ZRS失活,而蟒蛇仅约20%的序列差异给小鼠带来严重的四肢发育异常,让ZRS功能严重退化的关键因素一定藏在这一小部分序列中(这也是所有现存蛇类共同祖先都经历的改变)。
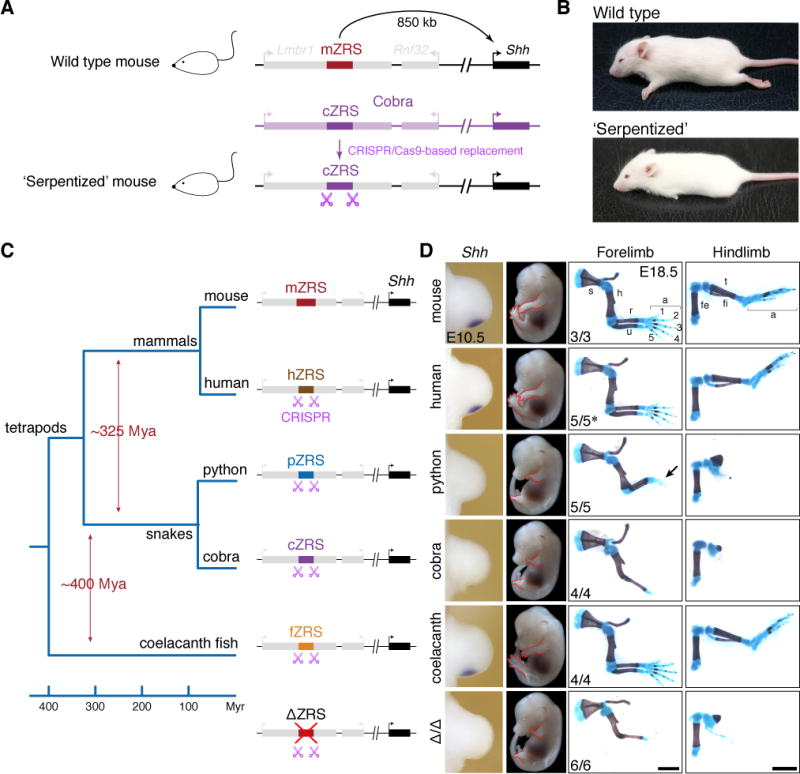
图2 不同物种ZRS增强子替代小鼠ZRS示意图,其中两种蛇类ZRS替换型小鼠与ZRS敲除型小鼠四肢发育严重受阻,它们的胚胎期检测不到Shh在肢芽特异性表达。
于是他们又进一步比对序列发现,所有蛇类都丢失了ZRS中17个碱基的核心序列(图3),而其他脊椎动物如人、鼠、鸡、蜥蜴、鲨鱼、腔棘鱼等覆盖有颌脊椎动物不同门类的基因组都包含这段序列,这表明虽然脊椎动物的四肢在形态与功能上各不相同,但发育出它们的上游调控机制是保守的,ZRS增强子出现至少可以追溯到有颌脊椎动物共同祖先的诞生阶段。此后绝大多数有颌脊椎动物的四肢都扮演着影响其生存方式的重要器官,故它们的ZRS增强子都呈现出极高的保守性;而蛇类则是在后来的演化历程中丢失了ZRS的17bp核心区域并因此几乎丧失四肢发育能力。尤其是对于那些彻底失去四肢发育能力的高级蛇类来说,ZRS已经是一个“死”的增强子元件,所以它们的ZRS可以不受进化约束,以一种“放飞自我”的方式呈现出超大幅度的突变。为了验证这个更进一步的猜想,Visel他们合成了一段补齐了这17个碱基的蟒蛇ZRS序列,再将其替换掉小鼠的ZRS,您猜怎么着?此时小鼠的四肢发育能力完全正常!它被“拯救”了出来!作为一个evo-devo科学家,Visel当然要从发育的角度解释清楚这中间发生了什么。他的团队先后在小鼠胚胎中做了不同的ZRS激活报告基因研究,结果表明:与其他ZRS拥有正常活性相反,替换到小鼠基因组中的蛇类ZRS失去了肢芽特异性激活的增强子特性,但补齐这17个碱基的蟒蛇ZRS则又获得了它的增强子功能,于是小鼠的四肢发育停滞被“治好了”。
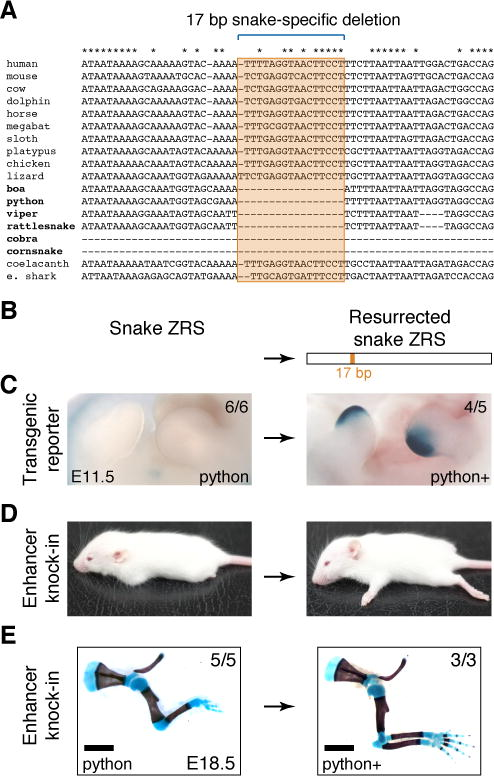
图3 蛇类ZRS增强子的17个碱基核心序列缺失以及补齐了这17个碱基的蟒蛇ZRS替换型小鼠重新恢复四肢发育能力。
讲到这里,我们能解答开头提出的问题了,假如我们建立了蛇类的基因编辑与转基因实验体系,补齐还没突变得面目全非的蟒蛇ZRS的这17个核心碱基之后极有可能让它们重新长出四肢。蛇失去四肢的案例是evo-devo理论的经典案例之一,我们接下来以ZRS-Shh调控关系为例看evo-devo理论如何填补经典演化理论的重大缺陷。
03. Evo-devo:生物学大一统理论的关键钥匙
1942年,英国生物学家J.S.Huxley在前人学说基础上提出了现代演化综论(Modern Synthesis, MS)[2]。这就是我们熟知的经典演化理论源,它来自对进化论和遗传学的综合,以种群内的基因频率变化为核心解释微观演化过程,而后又将宏观进化简单地解释为不断适应自然环境变化而产生的微观演化的累积。现代演化综论旨在凭借细胞学说、遗传法则等具有高普适性的基础理论来解释一切生物演化现象,力求争当生物学的“大一统理论”。而当雄心勃勃的生物学家们拿它去解释动物形态演化的时候,胚胎发育问题就成了他们的拦路虎。正如后来Evo-devo的主要提出者之一,Sean Carroll曾在他的《无尽之形最美》一书中指出的那样,“如果我们不能弄清某种动物如何发育成自身的成年形态,又怎么能知道它的祖先在形态演化的过程中取得了哪些进展?种群遗传学成功建立了基因变异导致演化的定律,但它没有任何实例…所以想要取得演化生物学新理论就必须在胚胎发育的研究上有所突破”[3](这本书是有关evo-devo最棒的科普读物,强力推荐)。
时间到了20世纪80年代,科学家们手持着分子生物学的武器重新征战古老的发育生物学领域,他们在果蝇体内找到了一系列控制果蝇发育的关键基因。而此时一场悄无声息的演化生物学革命正逐渐拉开了序幕。过去人们曾经想当然地认为,如果两种动物在形态上差别越大,它们共有的决定发育的基因就越少,例如MS的设计师之一,Ernst Mayr曾写道“除了非常接近的物种之间,寻找同源基因是没有意义的”。然而事实却恰恰相反,决定果蝇胚胎发育的大部分基因,大部分不仅存在于绝大多数动物中,甚至我们人类自身也有相应的同源基因(homologous genes),且这些基因在动物体内具有类似的功能。通过对来自不同物种发育基因的比较研究,在发育生物学和进化生物学的交叉中,进化发育生物学(evo-devo)就此诞生了。Evo-devo学派极力阐述将胚胎发育重新纳入演化理论体系中的必要性,我们认为:除了自然选择的外部因素对新物种形成起关键作用之外,发育调控关系作为复杂动物体内在因素对此同样起到关键作用。
与发育调控相关的蛋白总共分为两类:一类是在不同细胞之间信号传递着细胞增殖或分化信息的信号分子如上文提到的SHH蛋白;另一类是在细胞核内通过结合增强子而控制开启基因表达的蛋白,即转录因子,例如上文中提到的结合到ZRS那17个核心碱基上的ETS1蛋白。发育调控蛋白及对应基因在不同物种间高度保守,就以上文所讲的Shh基因为例,它广泛地存在于脊椎动物、节肢动物、乃至更早出现的棘皮动物的基因组中。而Shh能调控细胞分化,同时它也受其他蛋白的调控,当然这些蛋白的基因表达也是如此,这样多个级联的基因表达调控关系就形成了复杂的基因表达调控关系网络(gene regulatory network,GRN)。如图4所示,最早提出GRN概念的Eric Davidson等人把它描述成了一个多级互相作用的“电路图”[4]。
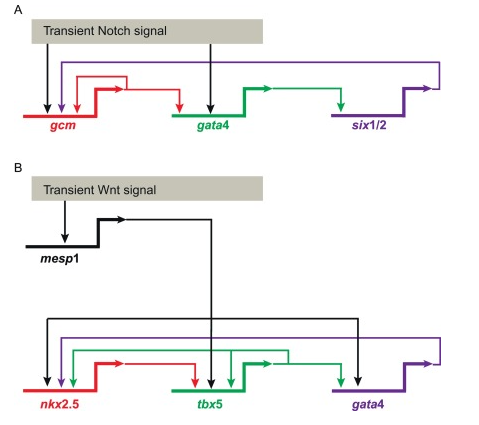
图4 某些器官发育的部分调控网络示意图
(A)Notch信号驱动海胆中胚层分化的调控网络
(B)Wnt信号驱动小鼠心脏分化的调控网络
讲到这里,一个深刻的问题浮出水面:之前当把肉鳍鱼的ZRS增强子替换给小鼠的时候,小鼠并没有长出鱼鳍而是依然发育出小鼠的四肢。ZRS-Shh调控关系对四肢发育的决定似乎是一条很高级又很模糊的指令,它只是要你发育出四肢,而具体长出什么样的四肢则取决于不同生物的执行方式,这就是evo-devo理论所指出的关键,调控网络的层级性。四肢发育的调控网络核心是在除蛇类之外的有颌脊椎动物中高度保守的,但后面能发育出什么具体形态的四肢则取决于下游的调控关系,不同物种在这里往往有复杂多变的调控关系所导致的不同四肢结构,例如鱼的鳍,鸟的翅,人的手。2006年,evo-devo学派领袖之一的Eric Davidson提出了本领域的几条基本原理[5]。他将复杂生物演化归因于无数新调控关系建立的过程,而那些处于上游的核心调控关系的建立则驱动了大的物种门类(如“超门”、“门”、“亚门”等结构)的保守特征的出现。如图5所示,虽然脊椎动物和昆虫的心脏结构相差甚远,但在它们早期发育阶段的核心调控关系确实高度保守的,这表明心脏为循环系统提供动力的功能可以追溯到的两侧对称动物的共同祖先,而跨越脊索动物、节肢动物等多个门的后生动物都在继承这种关键的发育调控关系的基础上发展演化。
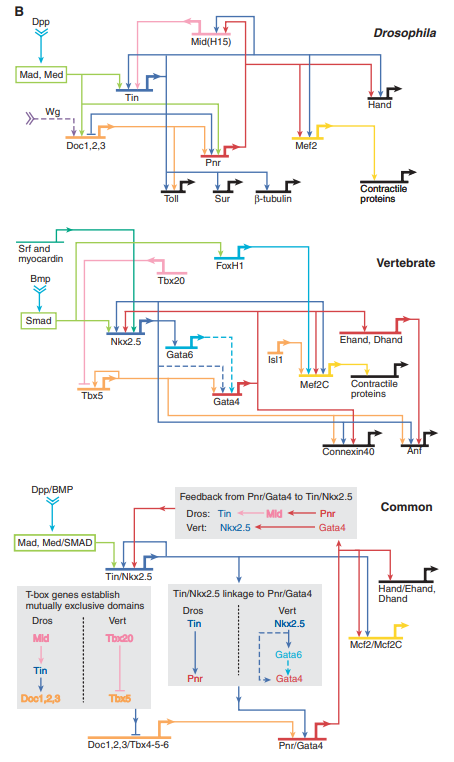
图5 果蝇与脊椎动物心脏发育特化过程的基因调控网络核心示意图,上图为果蝇,中图为脊椎动物,下图为二者共享的跨门类的高度保守调控关系。
Evo-devo所揭示的调控关系复杂性也并不仅体现在纵向的多层次结构,从横向层次看,复杂多细胞动物的基因组中,一个关键的发育调控基因往往也是受多个增强子调控以实现其在不同时空条件下激活表达,这使得它表达出的调控蛋白可以参与多个独立的发育过程。我们仍然以Shh为例,在脊椎动物中,Shh也参与了多个复杂的发育过程,例如神经系统,四肢,消化道,面部等多个生理组织或器官的发育过程。如图6所示,从基因组保守性的角度分析,我们会发现在Shh基因上游的巨大“基因沙漠”中不仅分布着一个结合转录因子ETS1的ZRS增强子,还有结合其他转录因子的增强子,它们参与着不同过程的调控,使Shh在胚胎的不同阶段不同位置表达。这些保守增强子构成了一个保守的基因组调控模块(genome regulatory block,GRB)[6],但它们彼此互不干扰,所以破坏了ZRS功能的小鼠只出现了四肢发育异常的情况,而没有影响到它的神经系统发育。这保证了Shh等多个关键发育基因既在上游受不同发育基因的调控而在不同位置表达,又在下游能激活不同基因的表达以实现不同的发育功能,都是堪比汤姆猫一样的“多面手”。虽然大部分关键的调控基因都可以追溯到原生动物甚至是真菌的基因组中,但是在演化历程中多层次的基因组调控关系的累积使得后生动物发育过程的复杂性呈现指数级爆发,由此产生了斑斓多样的外表形态。这就是Sean Carroll对“endless forms most beautiful”的诠释,无尽的多层次调控关系带来了生物的复杂之美。
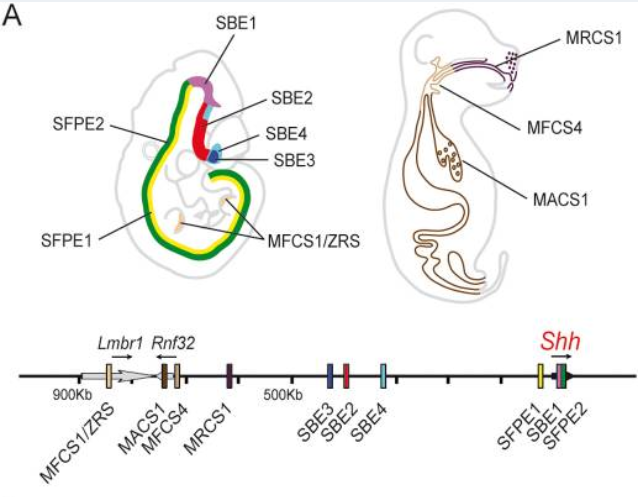
图6 组织特异性增强子在小鼠 5 号染色体中Shh的大上游区域和内含子的分布。每个增强子都表示为一个色块,其相关表达在上面的示意图胚胎中以相同的颜色显示。
基本概括了evo-devo的核心思想后,我们重新审视它如何为演化理论带来革命性的创新。在传统演化理论框架下,某个物种而言两个相对性状谁更有优势完全取决于环境差异,环境的改变完全可能逆转针对某个具体性状的演化方向,理论上微观演化过程是可逆的,而由无数微观演化累积而形成的宏观演化过程则是一个相对缓慢的渐变过程。然而那些带来物种爆发的关键演化节点的情况则无法用它来解释。我们人类演化过程历经了后生动物、两侧对称动物、后口动物、脊索动物、脊椎动物、有颌动物、四足动物、羊膜动物等几个决定我们今天生理形态的关键演化节点。它们出现的窗口相比于漫长的演化历程虽然十分短暂,但它们的出现都带来了极大规模的新物种形成,而且十分严格地决定了现生多细胞动物的基本发育框架。Evo-devo尤其关注这些带来物种迅速爆发的不可逆演化节点。我们认为:这些演化节点的出现背后是少数决定细胞模式多样性和生理结构复杂性的核心调控关系从无到有的突变过程,这是基于群体遗传学“渐变的微观演化累积”观点的传统演化理论所无法解释的。核心调控关系一旦建立,就会为后来的演化过程带来无尽的可能性和复杂性。例如在三胚层动物演化过程中,不管中胚层首次出现后被用于具体哪些细胞分化和器官发育,建立了分化出中胚层的调控关系本身就为三胚层生物带来了比二胚层生物至少多出一个数量级的分化潜能,所以这种进化创新过程整体上是高度不可逆的(不排除个别物种类群的局部发育过程因丢失一个核心调控关系而退化如蛇的四肢退化)。Evo-devo就是通过寻找涵盖某个关键演化节点的不同门类物种之间的基因组保守性差异来破解此节点对应的调控关系建立如何决定了物种演化的大方向。
我们都知道,几乎全部现生复杂动物门类的形成都能最早追溯到寒武纪时期,而寒武纪物种爆发则使进化论自达尔文时代就饱受创造论者的攻击,而秉持evo-devo观点的Eric Davidson则很容易地诠释寒武纪物种爆发过程:不同门的后生动物的GRN的“核心架构”最晚是在寒武纪前后搭建出来的(其中很多超保守的调控关系甚至跨门共享),而它一旦组装好,GRN上游的核心调控关系就不能随意拆卸或基本上重新布线,只能在此基础上继续搭建的下游调控关系。
04. 基因组上的一份神秘协议
互联网协议和通讯技术协议使得来自世界各地的信号在广阔的天地间交互而互不冲突。在单细胞形态停滞了数十亿年之后,一份有关规定了细胞间如何互相转化和相互作用的多细胞调控协议使得我们的祖先迈出了走向多细胞的最伟大一步。五亿多年以来,那最古老的祖先协议在我们所有后生动物的基因组里被抄送了无数份,而不同门类动物的基因组在此基础上又逐渐添加新的协议内容,赋予新的生命内涵,代代相传,它使我们一步步成为了今天的“人”。它既是个体发育的主函数,又是物种演化的日志文件,是铭刻在基因组上的演化史诗。它对多细胞动物至关重要,性命攸关,对它的探索必将带领我们一步步解开“我是谁,我从哪里来”这一终极谜题。
以上大概就是我决定投身于evo-devo研究的主要理由。(文末强烈安利我们进化发育之歌《evo-devo》,可以在B站搜得到)
参考文献
[1] Evgeny Z.Kvon, Olga K.Kamneva, Uirá S.Melo, et al. Progressive Loss of Function in a Limb Enhancer during Snake Evolution [J]. Cell, 2016, 167(3):633-642.
[2] Huxley J.S. Evolution, the modern synthesis [M].
[3] Sean B. Carroll. Endless Forms Most Beautiful [M].
[4] Isabelle S.Peter, Eric H.Davidson. Chapter Thirteen - Implications of Developmental Gene Regulatory Networks Inside and Outside Developmental Biology [J]. Current Topics in Developmental Biology, 2016, 117:237-251.
[5] Eric H. Davidson, Douglas H. Erwin. Gene Regulatory Networks and the Evolution of Animal Body Plans [J]. Science, 2006, 311(5762):796-800.
[6] Manuel Irimia, Jose L. Royo, Demian Burguera, et al. Comparative genomics of the Hedgehog loci in chordates and the origins of Shh regulatory novelties [J]. Scientific Reports, 2012, 2(433).








