水稻起源

说到主食,我们生活中最常见的要属稻米(Rice, Oryza spp.)。根据颜色,我们可以将稻米分为白米、红米、紫米、黑米;根据口感,我们有糙米、东北大米、泰国香米等;根据生态型,分为栽培稻、野生稻、杂草稻;根据育种方式,分成常规稻和杂交稻;根据种植条件,可分为水稻、陆稻、海水稻等等。还有一些,看着像米,但不一定是稻米,如市面上宣传具有很好保健功效的“野米”并不是来自野生稻米,而是来自菰属(Zizania)的菰米,江浙一带的食用蔬菜茭白的种子。

Fig1. 红米(杂草稻)与白米(栽培稻)
作为最重要的主粮和植物科学基础研究的模式物种,稻米如何起源及演化一直是被广泛讨论的话题。在种(Species)的水平上,目前学界并没有太大争议。世界上广泛种植的稻米有两个种,属单子叶植物纲禾本目禾本科稻属,分别为亚洲栽培稻(Asian cultivated rice, Oryza sativa L.)和非洲栽培稻(Oryza glaberrima Steud.)。前者主要种植在亚洲,驯化自普通野生稻(Oryza rufipogon),后者主要种植在非洲,驯化自短舌野生稻(Oryza barthii)。
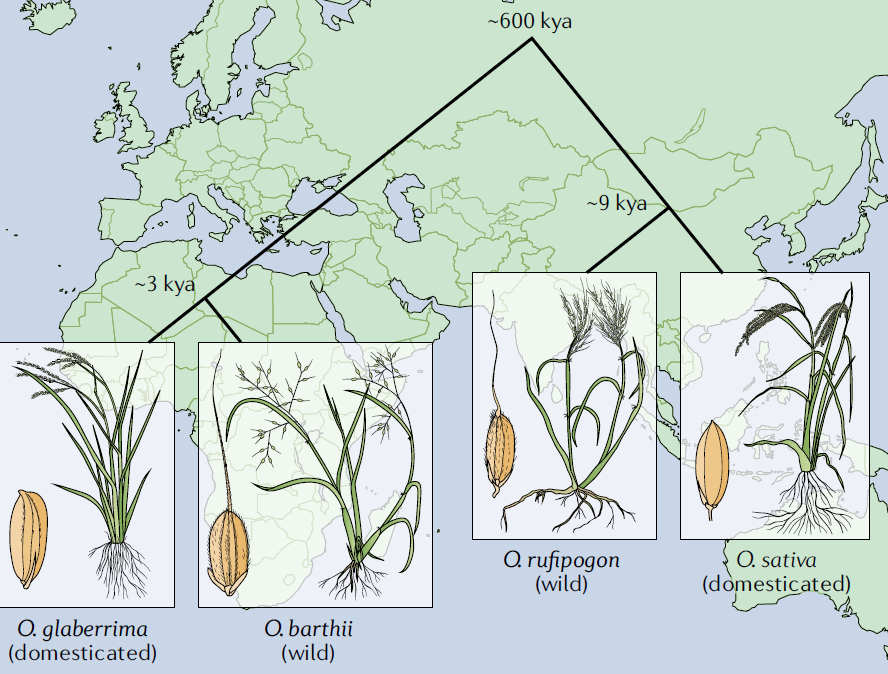
Fig2. 亚洲栽培稻和非洲栽培稻的起源与形态(Wing et al., 2019, Nat. Rev. Genet.)
从野生到作物,往往伴随一些明显的表型变化,统称为“驯化综合征(Domestication Syndrome)”,比如株型从匍匐变为直立(集约化种植)、谷粒落粒性降低(结实的种子掉了就没得吃)、种子休眠性降低(不容易发芽下一茬就没得种)等等,在此过程中介导表型变化的关键基因(驯化基因)也得到了广泛研究。
亚洲栽培稻的起源在哪里?
关于稻米起源,争议最大的是亚洲栽培稻的起源问题。种尺度下面又分为不同亚种(Subspecies)、亚种下面分为不同组(Group或者Clade)。一般认为亚洲栽培稻可分为两个亚种:籼稻(XI, ssp. Xian / indica)和粳稻(GJ, ssp. Geng / japonica)。籼稻主要种植在热带及亚热带地区,如我国南方、南亚、东南亚等,粳稻主要种植在温带地区及少数高海拔地区,如我国东北、日本、韩国等。籼稻又可划分为秋稻(aus/cA)、XI-2/、XI-3、XI-1A、XI-1B等组;粳稻可进一步分为温带粳稻(tmp,如东北大米)、热带粳稻(trp)、巴斯马蒂(Basmati/cB,俗称印度香米)等组。秋稻有时候也会当做是与两个亚种相当的分类等级。
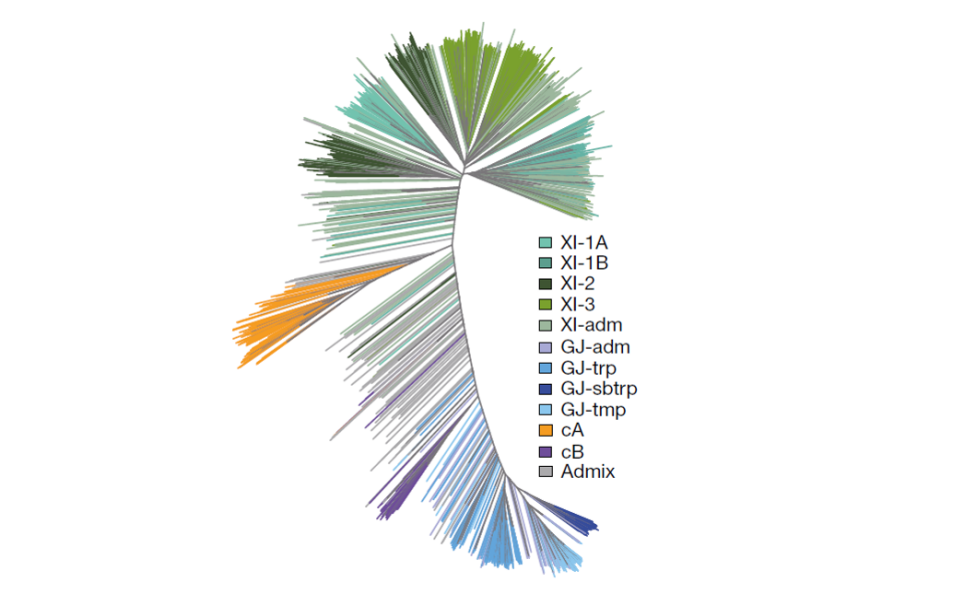
Fig3. 3K项目全球栽培水稻基因组的系统发育树(Wang et al., 2018, Nature)
根据“栽培植物起源中心学说”,20世纪初苏联遗传学家瓦维洛夫提出水稻起源于印度,原因是印度地区的稻属物种遗传多态性最高。后来在中国华南地区也发现野生稻,中国著名农学家丁颖先生在大量研究的基础上提出“水稻起源于中国华南地区”的观点。此外,喜马拉雅山南麓高物种多样性地带以及东南亚沿海沼泽栖息地也曾被认为是水稻起源地。
随着我国长江流域的很多新石器时代遗址中水稻遗存的陆续发现,著名考古学家严文明先生在20世纪90年代初提出“水稻起源于长江中下游地区”的观点。这其中代表性遗址之一是1973年发掘的、距今超过7000年的浙江余姚河姆渡遗址,由于积水缺氧的保存条件,遗址中的生物材料保存相对完好,这也为有效区分水稻遗存的驯化状态提供了条件。一些学者认为,河姆渡人可能有一个以水稻为基础的非常成熟的农业经济体系。此外,浙江上山遗址是目前所知长江下游地区最早的新石器时代遗址,最早年代可距今1.1万年,这一水稻遗存的发现轰动一时。虽然印度、泰国等地也曾陆续报道了距今8000年前的水稻遗存,但这些发现后来都被证明可靠性存疑。自此,“中国长江流域是水稻最初被驯化的地方”成为国际学术界的共识。
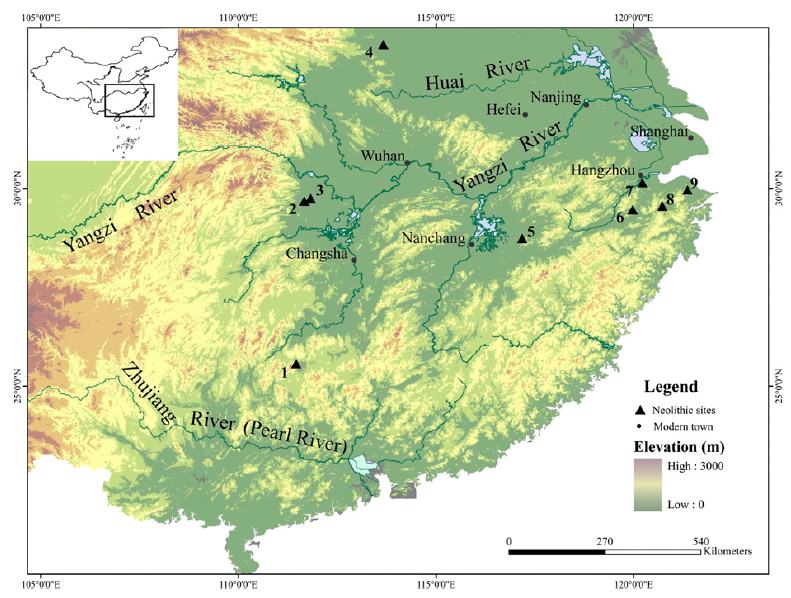
Fig4. 长江流域早期水稻遗存遗址(Gross & Zhao, 2018, PNAS)
野生材料的有无对于判断起源地很重要。目前,亚洲栽培稻的直接祖先野生稻O. rufipogon(普通野生稻)主要分布在南亚、东南亚和我国华南地区(北至江西一带,如东乡野生稻),且不同地区的野生稻存在显著的遗传差异和群体结构。比如我国华南地区的野生稻主要类群是Or-III,南亚主要是其他三种(Or-I/Or-II/Or-IV)。Or-IV是普通野生稻的基部类群,因此可以认为南亚地区是野生稻的起源地,中国华南地区的野生稻Or-III群体演化自南亚地区的祖先群体。
水稻多样化类群怎么形成的?
我们已经知道了最早驯化水稻的地方,但目前多样化的水稻类群是如何形成的呢?籼稻和粳稻在表型上具有很多差异,比如籼稻籽粒更加修长,而粳稻更加圆润;籼稻耐热不耐冷、粳稻更耐冷等。尽管这些性状的差异一部分是由于自然条件的局部适应性所致,但是籼粳稻如何演化,并产生当下分布格局,依旧是个有趣的问题。学术界有两种主流假说:单次驯化多次起源假说(Single domestication and multiple origins),和多次驯化(Multiple domestication)假说。即籼稻和粳稻是一次驯化后分散到各个地区,随后经历了漫长局部适应(包括自然环境和人类选择)形成表型上的分化?还是各自独立驯化自不同的野生稻群体?

Fig5. 野生稻(左)与栽培稻(右)(Tan et al., 2008, Nat. Genet.)
驯化基因或许能直接反映两个栽培亚种的关系。对控制落粒性关键基因sh4的分析表明,控制非落粒表型的突变等位基因属于单一起源,单系涵盖栽培粳稻和籼稻(Sang & Ge, 2007, Curr. Opin. Genet. Dev.; Zhang et al., 2009, New Phytol.)。这一重要驯化特征背后的基因单一起源促使研究人员考虑“互动驯化interactive domestication”场景的可能性,如中科院植物所葛颂研究员提出的“滚雪球模型”和“组合模型”(Sang & Ge, 2007, JIPB)。这些模型开始尝试通过引入基因流的各种场景来协调亚种间中性基因座的差异性与驯化基因座的相似性。
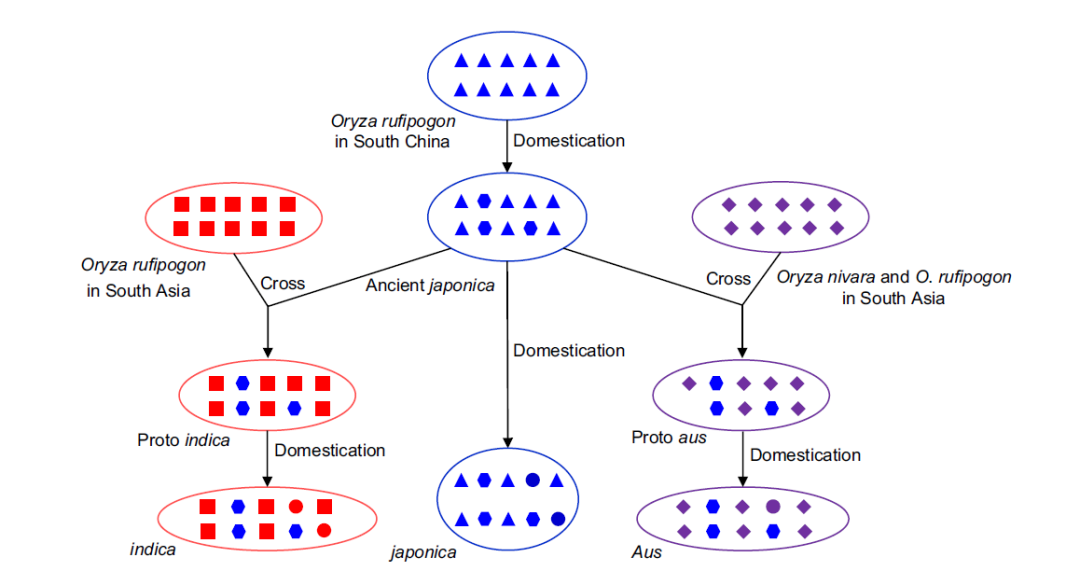
Fig6. 遗传渗入对水稻栽培亚种形成的影响过程(Wei & Huang, 2019)
基于分子标记和驯化基因上的变异信息终究有限。2012年,中科院分子植物卓越创新中心韩斌院士团队完成了446个的野生稻基因组和1083个籼粳基因组重测序的分析工作(Huang et al., 2012, Nature),全基因组变异特征和对选择性清除(Selective Sweep)区域的分析表明,粳稻与中国华南地区(珠江流域而非长江流域)的普通野生稻之间有着更加密切的关系,同时也强烈支持“一次驯化与多次起源”观点。随后在2015年,来自英国的Terence A. Brown团队发文质疑Nature文章的方法(Civán et al., 2015, Nat. Plants),并基于相同数据集和新的分析策略提出“栽培稻不同亚种是独立驯化而来”的观点。随后两个团队进行了多次回应(Huang & Han, 2015, Nat. Plants; Civán et al., 2016, Nat. Plants)。2018年,纽约大学Michael D. Purugganan团队利用同一批数据分析认为,不同亚种起源于多个野生稻群体,这一点从全基因组水平的系统发育关系可以清晰看到,但从头驯化事件仅发生过一次,且发生在粳稻中,籼稻的驯化等位基因来源于亚种间遗传渗入(Choi & Purugganan, 2018, G3)。
基于比较基因组学和溯组模拟分析也支持上述结论(Choi et al., 2017, MBE)。随后,更多类型的变异数据、分析方法被应用到起源推断上。如基于转座子插入多态性(Transposon Insertion Polymorphism)的研究认为栽培稻不同亚种来源于独立的驯化事件(Carpentier et al., 2019, Nat. Commun.)(Nat. Commun. | 利用3K水稻数据研究水稻还原转座子进化)。基于编码区域变异位点的单倍型(gcHap)分析认为水稻驯化是一个multiorigin (domestication) model (Zhang et al., 2021, Molecular Plant)。整体来说,尽管有些研究认为水稻驯化是多次的,但仔细分析结果就会发现这样的推论没有足够的证据支持。
因此,目前越来越多的学者支持“一次驯化和多次起源”假说,即粳稻首先在中国南方驯化自当地的普通野生稻(Or-III),随后伴随贸易等人工活动传播至南亚地区,很可能与当地的野生稻(Or-I)发生杂交和遗传渗入,这里我们无法排除渗入受体是古籼稻(经历了初步或正在经历驯化的群体)的可能性,尽管考古学和遗传学都没有明确提供南亚当地已经开展水稻初步驯化的证据。重要驯化等位基因(落粒性sh4、芒长an-1/LABA1、种皮颜色Rc等)被选择性保留在古籼稻群体中,随后籼稻、粳稻各自经历了多样化(Diversification)和育种改良(Improvement)过程,最终形成当下的地理分布格局和遗传特征。
至于亚种下的更小分类级别的不同水稻群体是如何分化、形成和传播的,可以参考Michael D. Purugganan团队利用基因组、考古、古环境/气候等综合建模的分析(Gutaker et al., 2020, Nat. Plants)(Nat. Plants | 重磅!最专业详实的水稻起源传播基因组证据)。如,既然粳稻最早在中国南方被驯化,那中国南方现在的籼稻是哪里来的?建模结果表明,可能是完成驯化的印度籼稻通过“丝绸之路”或横断山脉传播来的。传播时间与佛教书籍记录相吻合,有关佛教从印度传入中国的时间大约为1950年前。印度和中国亚群之间的密切关系反映在两个地区较高比例的灌溉品种上。相比之下,东南亚品种多进行雨养。
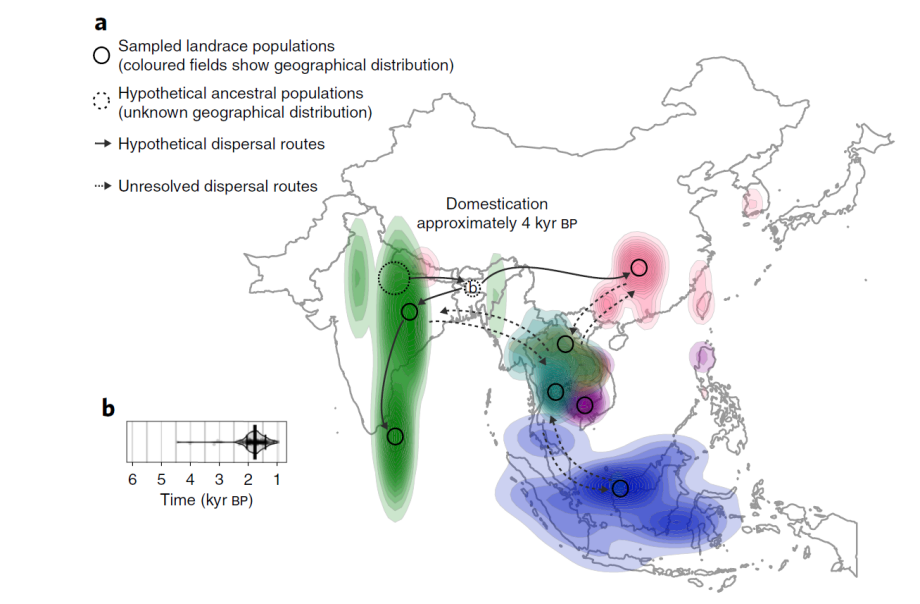
Fig7. 籼稻在亚洲的传播地图(Gutaker et al., 2020, Nat. Plants)
基于分子数据推断过去的演化历史的一个常用原则是最大简约。一个性状从形成到被固定在一个群体、一个物种中需要万年甚至更久的时间。驯化是一个缓慢的过程,如果有现成可用的驯化等位基因可以用,那会大幅降低演化成本。粳稻的遗传多态性远低于籼稻(暂且忽略南亚地区野生稻对现代栽培籼稻的遗传渗入),亚种间多态性的差异也许能够反映两者驯化历史的绝对差异。场景回到驯化初期,先民不可能只钟情于稻子,其他野草也会被种植,只有那些具有有利性状的材料才会被保留下来,这样的材料屈指可数,因此初期的选择瓶颈应当是非常大的。对于籼稻来说,高遗传多态性一定程度上意味着独立被驯化的可能性不大。
对于遗传渗入的思考
遗传渗入在作物驯化的过程中非常普遍。与自然群体不同,作物与人类的互作克服了地理隔离,加速了遗传变异在不同群体、不同亚种甚至是不同物种之间的传播。比如,现代六倍体栽培小麦的驯化等位基因是从不同地区的地方小麦群体(包括不同基因组倍性)的不同驯化基因聚合而成(Wang et al., 2022, Nat. Commun.)。
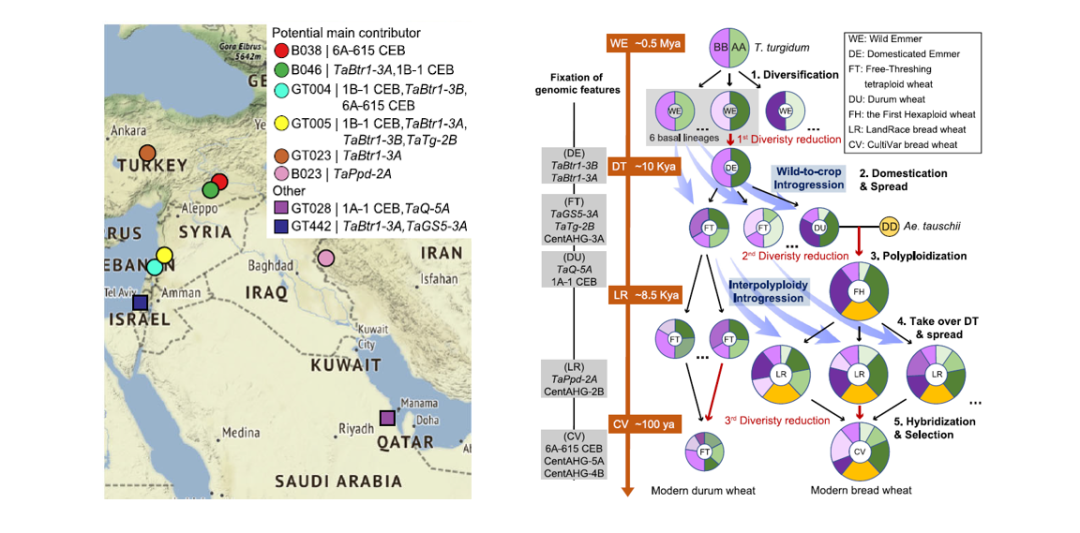
Fig8. 六倍体栽培小麦的驯化基因多起源和逐步聚合模型(Wang et al., 2022, Nat. Commun.)
它山之石,可以攻玉。遗传渗入在塑造物种多样性上的作用可能远比我们了解的大。遗传变异的产生会导致表型的多样化,我们可以经常在相近物种里看到类似的表型多样性,这其中多少是独立的趋同演化?多少是相互的基因渐渗?我们现在所观察到的物种间的生殖隔离在万年甚至是百万年以前可能都并不存在。
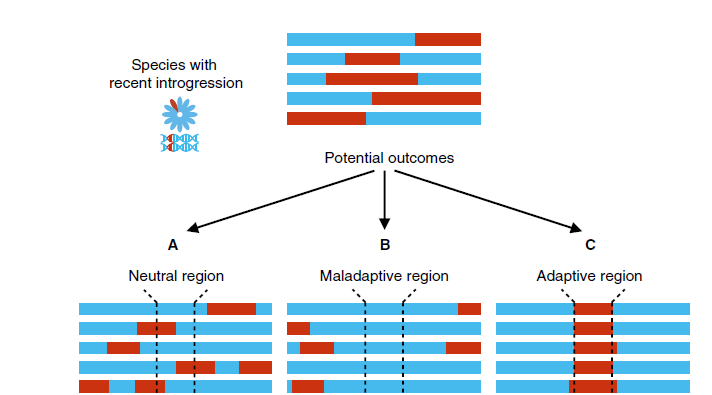
Fig9. 初始渗入事件后的潜在结果(Aguillon et al., 2022, Curr. Biol.)
在强的选择压力下,遗传渗入(如果有近缘等位基因的存在)比自发突变可能更容易发生和保留,并且被保留下来的遗传渐渗片段一定在适合度上有利自然选择(Natural Selection)或符合选择目标(Artificial Selection),尽管可能会存在一定程度的生殖隔离。








