EES讨论第一期|扩展演化综论——关于演化的争论波涛再起
2022年6月28日《卫报》的一篇名为「Do we need a new theory of evolution? 」的报道[1],将关于演化论的讨论推向了舆论的风口浪尖。中文互联网社区也有了相关的转载,并极具煽动性地以“我们是否需要一种新的进化论?”为标题,传播得极为广泛,评论区的讨论尤为热烈。

朱利安·赫胥黎和其所著的现代演化综论
文章中所要“推翻”的演化论,是被演化生物学家广泛接受的传统演化论(Standard evolutionary theory, SET)。传统演化论以现代演化综论(Modern Synthesis, MS)为核心,理论以基因为中心,将改变基因频率的事件视为演化过程。
现代演化综论(MS)出现在二十世纪上半叶,由演化生物学家Julian Huxley整理融合了当时达尔文的自然选择,群体遗传学和孟德尔遗传等理论而成,该理论为演化生物学提供了主要的概念框架[2]。在传统的演化论中,认为影响演化的四大因素分别为:突变(Mutation),自然选择(Natural selection),遗传漂变(Genetic drift)和基因流动(Gene flow)
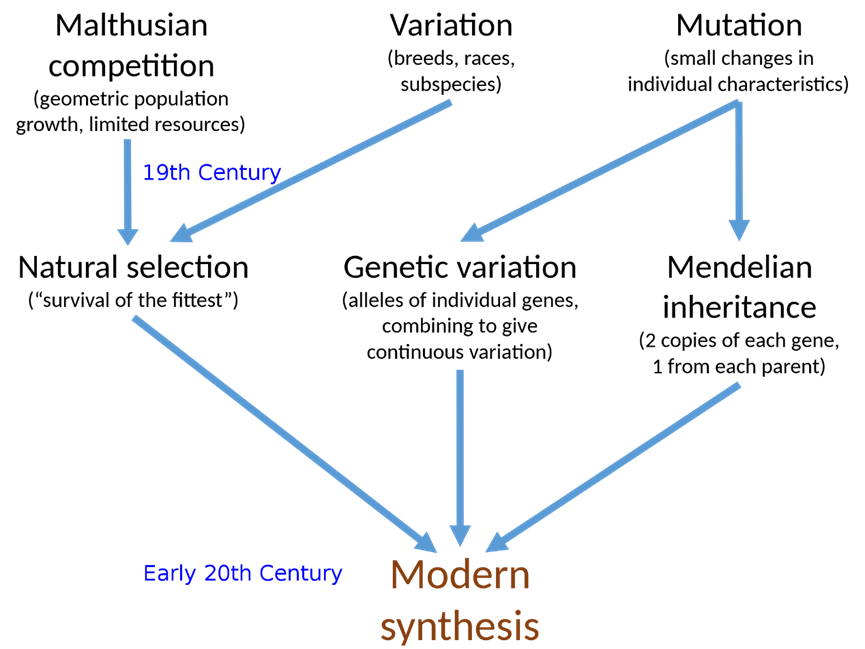
现代演化综论(MS)的主要内容和出现过程
近几十年来,以现代演化综论(MS)为核心的传统演化论(SET)指引了演化生物学的快速发展,取得了巨大的成功。然而,近年来越来越多生物学领域的新发现促使部分学者认为传统演化论需要补充新的内容并纠正一些缺陷。这些领域包括社会生物学、发育生物学、表观遗传学、微生物学、基因组学、水平基因转移等。
2007年纽约市立大学研究生中心的Massimo Pigliucci首次提出了对现代演化综论(MS)进行扩展的想法。他重新审视了传统演化论(SET)中包含和不包含的各种要素与它们之间的相对重要性,认为现代演化综论(MS)并不完善,一些新的要素需要被添加进演化理论,例如表型可塑性,可演化性,表观遗传和生态位建设等等[3]。2010年,他又与维也纳大学的Gerd B. Müller一起出版了《演化,扩展综论》(Evolution, the Extended Synthesis)一书[4],该书如今被认为是扩展演化综论(EES)的起点。

《演化,扩展综论》及两位作者
目前,扩展演化综论(EES)的支持者们已经组建了一个跨国跨学科的联盟。该联盟包括来自八个顶尖大学的51名世界知名专家,由圣安德鲁大学的Kevin Laland教授和隆德大学的Tobias Uller教授领导。关于该联盟组建的目的,Kevin Laland说到:“我们并不寻求用扩展演化综论(EES)取代传统的演化理论,而是希望EES能与SET共存,一起促进演化生物学的发展。”
目前,扩展演化综论(EES)的支持者们将其重新梳理为两个核心思想和四个焦点话题。
1. 扩展演化综论的核心思想
C1 构造性发育
constructive development
SET一般认为发育过程严格按照遗传程序进行的,发育过程中对环境的反应也是被预先设计好的。这种对于发育的认识有时被称为程序化发育(programmed development)[5]。EES支持者们所认为的发育过程是更加不确定的,更加积极的。EES认为:虽然,发育过程十分依赖遗传信息,但不完全由其决定;生物体在发育过程中会不断响应外界环境变化,同时也不断反馈甚至改变外部环境。这是一个探索性的过程[6]。
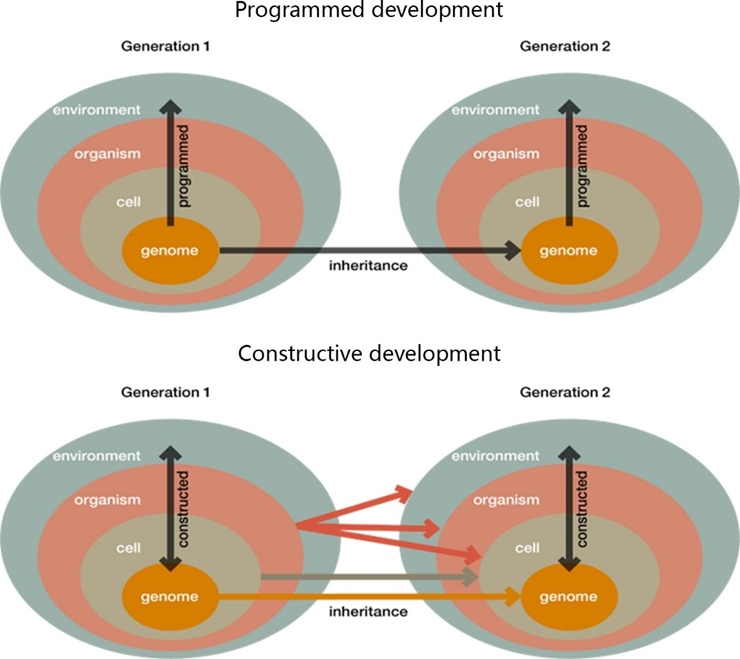
程序化发育(programmed development)和构造性发育(constructive development)的模式图
例如,循环系统神经系统的发育很好地体现了发育过程的探索性。循环系统的形状是根据各组织对氧气的需求构建的而不是遗传信息预先确定的。同样,神经系统的发育依赖于轴突的探索。肌肉纤维最开始总是能接触到多个神经元的突触,随后的突触竞争过程会加强某些神经元的连接而弱化其他的。这个过程中充满了不确定性,是无法预先被遗传确定的,但却决定了肌肉神经的最终形态和分布。
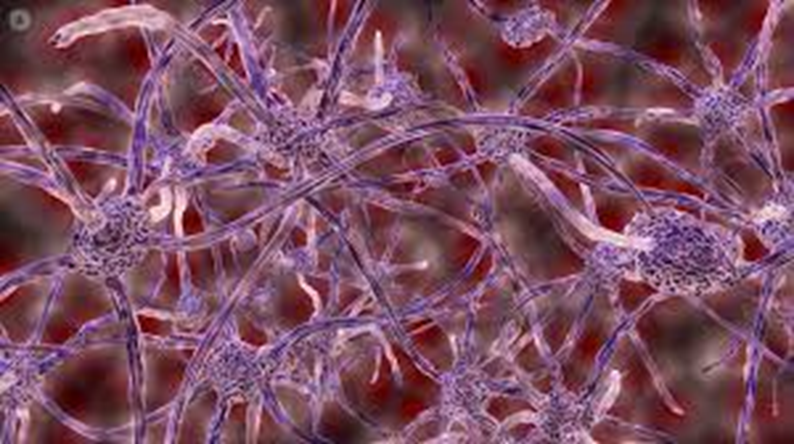
肌肉组织内的神经纤维
发育过程的积极性和探索性对于生命演化来说是非常重要的,具有建设性的意义。因为构造性发育可以根据遗传或者环境变化不断出现新的表型,这些表型将成为自然选择的新对象,并可能促进适应和多样化过程。
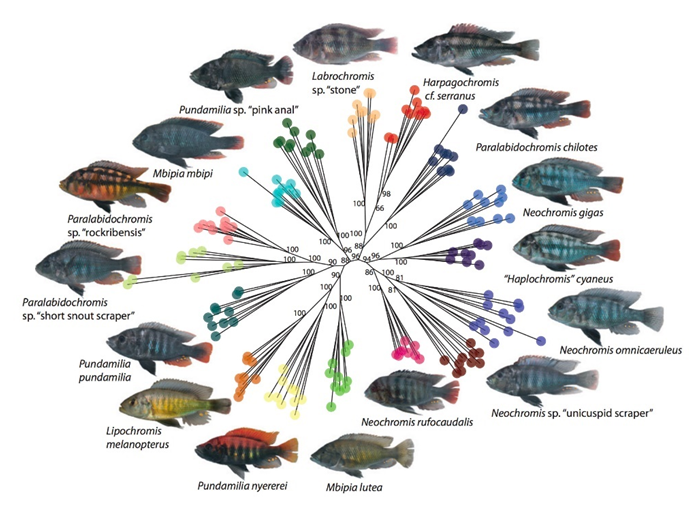
三湖慈鲷的构造性发育导致了它们变化无穷的表型和典型的适应性辐射[7]
C2 相互因果关系
Reciprocal causation
SET的观点认为生物演化的唯一原因是来自环境的自然选择。而生物发育过程中对环境的响应(发育可塑性)以及对环境的改变(生态位建设)都被视为自然选择作用下的不断循环的响应过程。而EES认为这些过程都是驱动生物演化过程的原因,与自然选择具有同等重要的地位。因此,发育中的生物体不仅仅是演化的产物,也是演化的原因,即生物-环境系统中的因果关系本质上是相互的。
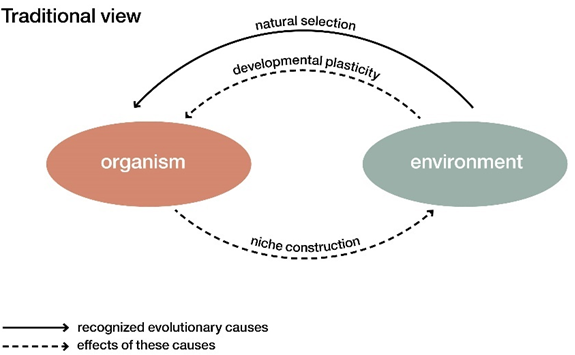
传统演化论(SET)和扩展演化综论(EES)关于生物与环境的关系示意图
2. 拓展演化综论的焦点话题
F1 发育偏好性
Developmental Bias
按照传统的演化理论,由于基因突变是随机发生的,因此性状变异也不会偏好任何特定方向。然而,许多生物学家怀疑,表型变异可能并不随机。某些特征总是比其他特征更容易、更频繁地出现。这被称为发育的偏好性(Developmental Bias)。
非洲坦噶尼喀湖和马拉维湖的慈鲷可以很好地阐述这一概念。慈鲷在两个湖泊内分别独立地发生了适应性辐射演化。虽然同一湖泊内的物种遗传上密切相关,但每个物种均在另一湖泊内存在表型上十分相似的物种(平行演化的大规模发生)。值得注意的是,其中有些性状比其他性状更具有可塑性,例如不同的下颌形状和牙齿形态将有利于对不同的栖息地和食物的利用。这一例子表明慈鲷可能特别擅长利用生态环境产生新的变异,从而导致物种形成和适应演化。此外,有些表型出现的难度明显高于其他表型,这种发育偏好会限制这些表型的演化。例如两个湖中的慈鲷都没有演化出具有夜行性、鳗体型或者掠食性的鱼类。

非洲坦噶尼喀湖和马拉维湖内慈鲷的表型多样性[8]
在偏瞳蔽眼蝶(Bicyclus anynana)翅膀上眼点的大小和颜色的拮抗选择现象,也可以进一步证明发育偏好性的存在:对于眼点一大一小的选择很快就能产生效果,然而一金一黑的选择却很难出现[9]。显然,发育过程中对于不同选择的响应速度和强度也是不同的。此外,诸如哺乳动物牙齿的数量和形态、动物四肢、手指和躯干的节段数量等都体现了发育偏好性的在自然界的广泛存在。
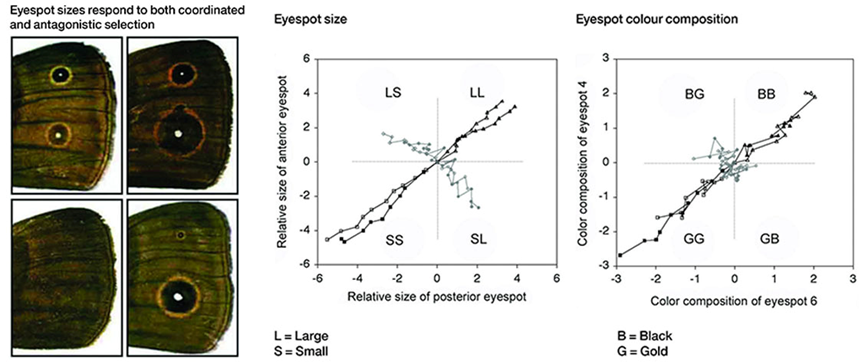
偏瞳蔽眼蝶翅膀眼点的大小和颜色的拮抗选择
F2 发育可塑性
Developmental Plasticity
发育可塑性是生物在环境条件变化时改变表型的能力,可塑性通过形态和行为的补偿性调整,帮助发育过程在面对环境波动时的稳健性。近年来,人们越来越多地认识到发育可塑性在演化过程中的作用。例如,促进物种产生、适应性辐射、并解释适应峰位移。
有科学家将发育过程在演化过程中的作用比作隐性遗传变异(cryptic genetic variation)的电容(隐性遗传变异指的是不会改变表型的变异)。正常情况下,生物的发育过程中会不断积累隐性变异,这也是积累发育可塑性的过程。当环境发生变化时,这些隐性变异就会表现出来,就像是电容释放电荷的过程。上面这种环境诱导的新表型会在几个时代的时间内微调并稳定下来,例如暴露在高捕食者压力下的水蚤会发生颜色丢失,颜色丢失一开始是具有可塑性的,然后会逐渐固定下来[10]。这个过程称为遗传顺应(genetic accommodation),是促进快速演化的重要机制。
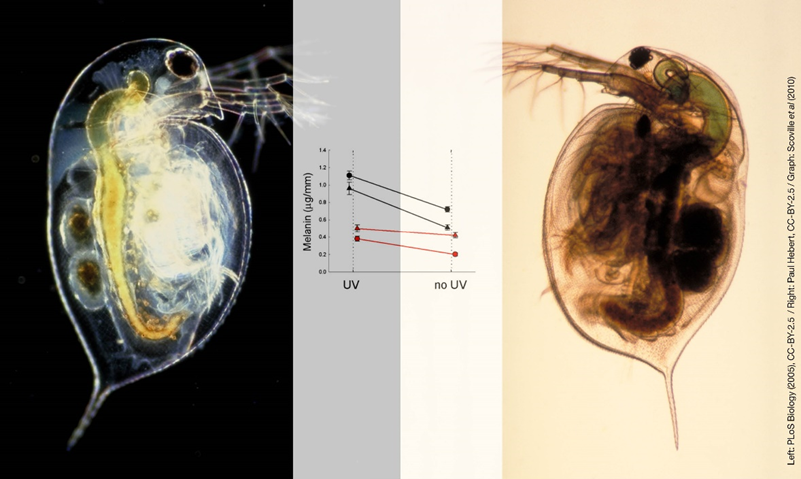
捕食者暴露下水蚤表型的发育可塑性和环境适应[10]
后来科学家进一步认为祖先物种的发育可塑性可能是后代物种性状多样性的起源,这被称为“柔性茎(flexible stem)”假说。
加拉帕斯雀可能是支持这个假说的一个例子。这种雀非常善于探索和占有新的生态位,或者说具有高度创新性。它们也因此具有很高的表型多样性,这被认为是来源于祖先种群的高发育可塑性[11]。

加拉帕斯雀的表型多样性[11]
鱼类和两栖动物中的例子进一步证明,某些生物类群的高物种多样性与发育过程中的高可塑性有关[12]。
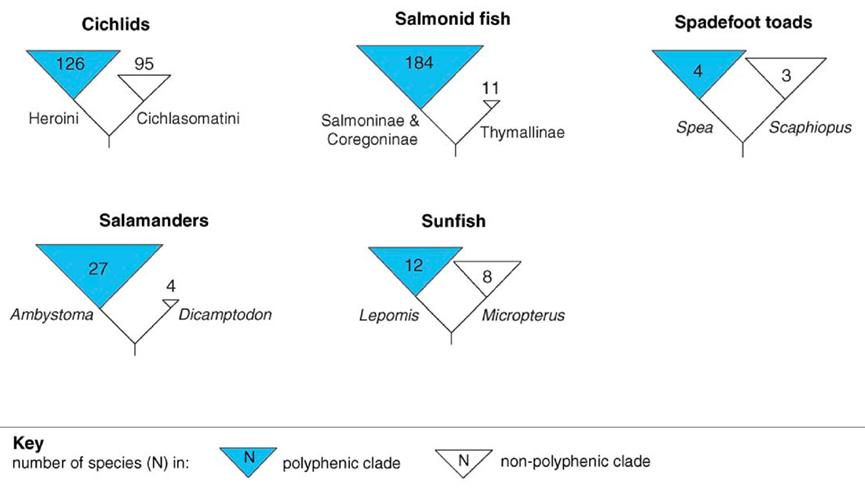
鱼类和两栖动物中某些类群的高物种多样性与发育过程中的高可塑性有关[12]
F3 包容性遗传
Inclusive Inheritance
除了传统意义上的遗传物质也就是基因或者DNA以外,还有其他的生物因子也是可以跨世代传递的,这些因子的传递可能是持续在整个发育过程中的,不仅仅只有通过生殖细胞传递这一种方式。这种观点被概括为包容性遗传(Inclusive Inheritance)[13]。
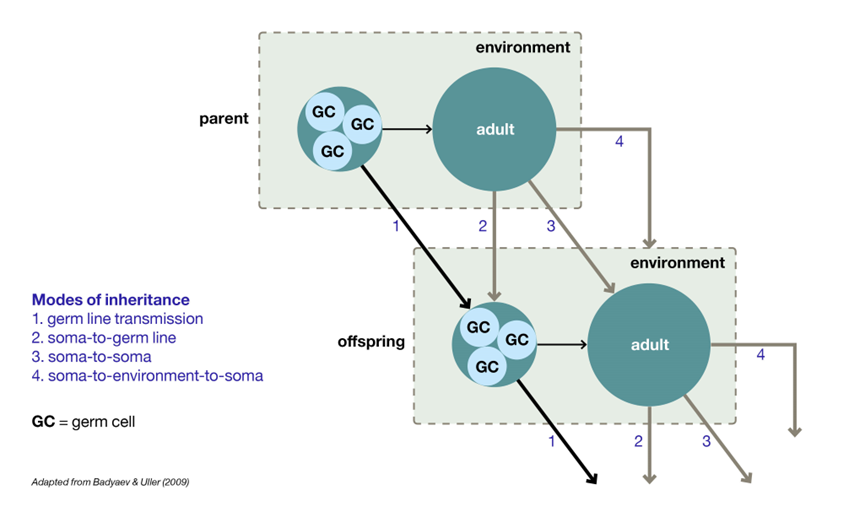
物质和信息可以通过生殖细胞之外的途径遗传和继承[14]
首先通过受精卵传递的不仅仅是DNA,例如细胞器、酶、激素、抗体和转录因子都可以通过受精卵传递给发育中的胚胎,并影响后代的表型,甚至成为受到选择的新变异的来源。一些表观修饰化学物质也可以附着在DNA或者染色体上跨代传递,例如DNA和组蛋白甲基化乙酰化修饰基团等。

各种各样的蛋
此外,来源于母体的微生物组可以主动或者被动地传递给后代,影响后代地发育、形态和抵抗力。例如:蜣螂育雏球内的母体微生物组对于幼虫地正常发育非常重要,特别是外界环境比较恶劣地情况下[15]。包容性遗传还包含文化传承。文化传承指的是通过交流、模仿、教学和学习来存储和传递非物质信息。它发生在从蚂蚁到人类的许多动物身上[16]。生态遗传也被认为是包容性遗传的一种。有些物种的亲代会改造生活环境并将其传递给后代。例如寄居蟹居住的贝壳大多数情况下都是从亲代那里继承来的,并且已经经过了亲代的改造[17]。

包容性继承的典型例子——蜣螂(母体微生物)、碎米荠(表观修饰)艾草榛鸡(学习行为)和生态遗传(寄居蟹)
F4 生态位建设
Niche construction
几乎所有生物都有积极地参与所在生态位的构建的特性[18]。比较著名的例子包括,海狸会在河里筑起水坝、沙漠中的自灌溉植物可以创造一个迷你绿洲、珊瑚礁为各种各样的海洋生物提供栖息地,还有蓝藻这样用数百万年改造了整个地球生态环境的极端例子。

生态位建设的典型例子——海狸建设的水坝(左上)、自灌溉的沙漠大黄(右上)、珊瑚礁生态系统(左下)和蓝藻改造早期地球生态系统(右下)
生物的生态位建设反过来对生物造成选择压力的改变。因此生态位建设和自然选择之间存在反馈关系。适应是一个双向过程。
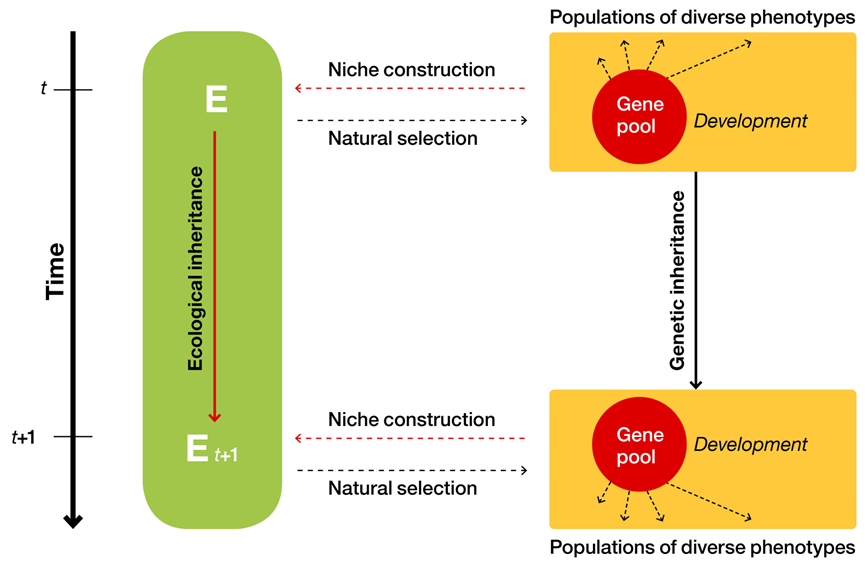
EES理论体系中环境和生物演化之间的关系
写在最后
SET认为表型的演化是在突变、选择、漂变和基因流的驱动和影响下进行的。而EES认为发育偏好性和生态位建设先于突变、选择、漂变发挥作用,并影响它们的作用,因而可以引导演化的方向。
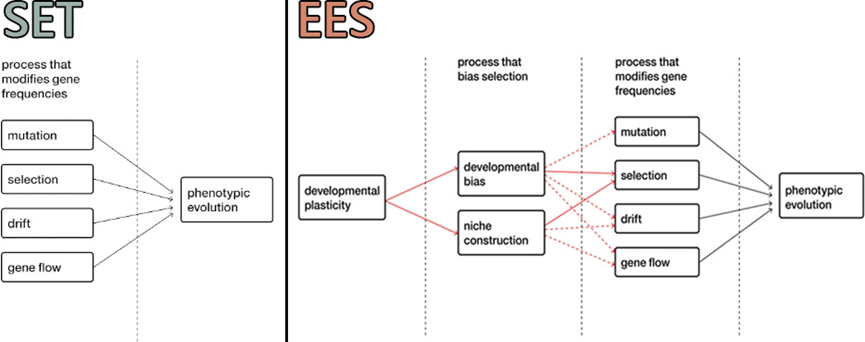
SET和EES对表型演化过程的理解存在区别
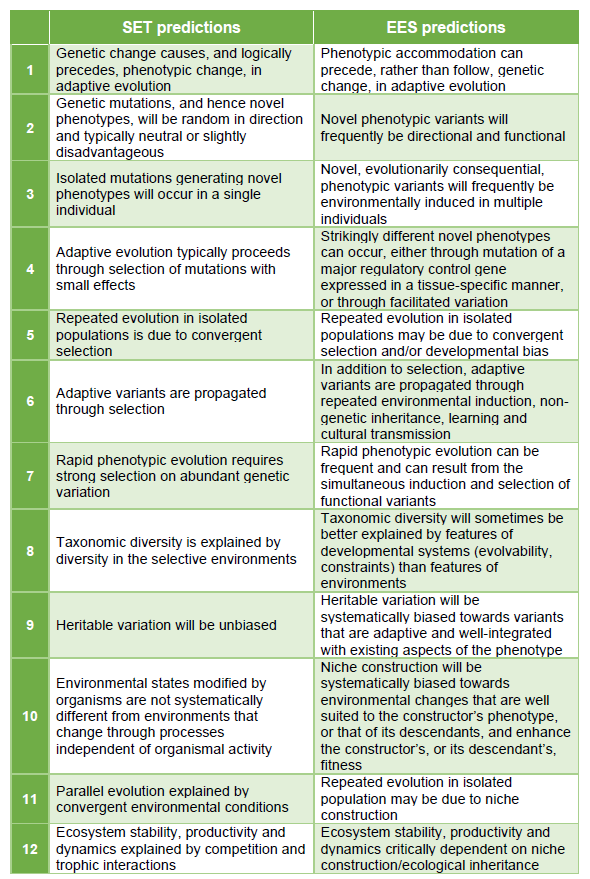
围绕着两个核心思想和四个焦点话题,EES的支持者们总结出了12条与SET相悖的对演化现象和过程的预测。限于篇幅原因,本文不对它们进行一一介绍。生命演化研究中心将组织四至五次关于EES和SET核心区别的讨论会,会上的重要内容也会以科普文章的形式在本公众号上发布,欢迎感兴趣的读者与我们积极讨论,我们会仔细阅读回复每一条留言。
扩展演化综论(EES)与传统演化论(SET)的主要区别
撰稿/陈葆华&陈光霁
编辑/刘磊
参考文献
[1] BURANYI S. Do we need a new theory of evolution? [N]. The Guardian, 2022-2022-06-28.
[2] HUXLEY J. Evolution. The modern synthesis [J]. Evolution The Modern Synthesis, 1942.
[3] PIGLIUCCI M. Do we need an extended evolutionary synthesis? [J]. Evolution: International Journal of Organic Evolution, 2007, 61(12): 2743-9.
[4] PIGLIUCCI M, MULLER G B. Evolution–the extended synthesis [J]. 2010.
[5] CASCIO S M, WASSARMAN P M. Program of Early Development in the Mammal - Post-Transcriptional Control of a Class of Proteins Synthesized by Mouse Oocytes and Early Embryos [J]. Dev Biol, 1982, 89(2): 397-408.
[6] LALAND K N, ULLER T, FELDMAN M W, et al. The extended evolutionary synthesis: its structure, assumptions and predictions [J]. Proceedings of the royal society B: biological sciences, 2015, 282(1813): 20151019.
[7] WAGNER C E, KELLER I, WITTWER S, et al. Genome‐wide RAD sequence data provide unprecedented resolution of species boundaries and relationships in the L ake V ictoria cichlid adaptive radiation [J]. Molecular ecology, 2013, 22(3): 787-98.
[8] ALBERTSON R C, KOCHER T D. Genetic and developmental basis of cichlid trophic diversity [J]. Heredity, 2006, 97(3): 211-21.
[9] ALLEN C E, BELDADE P, ZWAAN B J, et al. Differences in the selection response of serially repeated color pattern characters: Standing variation, development, and evolution [J]. BMC Evolutionary Biology, 2008, 8(1): 94.
[10] TOLLRIAN R, HEIBL C. Phenotypic plasticity in pigmentation in Daphnia induced by UV radiation and fish kairomones [J]. Funct Ecol, 2004, 18(4): 497-502.
[11] TEBBICH S, STERELNY K, TESCHKE I. The tale of the finch: adaptive radiation and behavioural flexibility [J]. Philos Trans R Soc Lond B Biol Sci, 2010, 365(1543): 1099-109.
[12] PFENNIG D W, MCGEE M. Resource polyphenism increases species richness: a test of the hypothesis [J]. Philos Trans R Soc Lond B Biol Sci, 2010, 365(1540): 577-91.
[13] DANCHIN É, CHARMANTIER A, CHAMPAGNE F A, et al. Beyond DNA: integrating inclusive inheritance into an extended theory of evolution [J]. Nature Reviews Genetics, 2011, 12(7): 475-86.
[14] BADYAEV A V, ULLER T. Parental effects in ecology and evolution: mechanisms, processes and implications [J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2009, 364(1520): 1169-77.
[15] PARKER E S, DURY G J, MOCZEK A P. Transgenerational developmental effects of species‐specific, maternally transmitted microbiota in Onthophagus dung beetles [J]. Ecological Entomology, 2019, 44(2): 274-82.
[16] JABLONKA E, SZATHMáRY E. The evolution of information storage and heredity [J]. Trends Ecol Evol, 1995, 10(5): 206-11.
[17] LAIDRE M E. Wild vs. Lab Box 10.2 Social Cognition in the Wild: From Lab to Field in Hermit Crabs [J]. Field and laboratory methods in animal cognition: a comparative guide, 2018: 237.
[18] LALAND K, MATTHEWS B, FELDMAN M W. An introduction to niche construction theory [J]. Evol Ecol, 2016, 30: 191-202.








