真菌王国的多样性与演化
真菌是一类真核生物,它们的细胞含有膜结合的细胞器和清晰的细胞核。真菌王国中预计有2~5百万种物种,目前大约有14.4万种已知物种,包括酵母、锈菌、黑穗病、霉菌、霉菌和蘑菇[1]。还有许多类真菌的生物,包括黏菌和卵菌(水霉菌),它们不属于真菌界,但通常被称为真菌。许多类似真菌的生物都属于变色菌属。真菌是地球上分布最广泛的生物之一,具有重要的环境和医学意义。许多真菌在土壤或水中自由生活,另一些则与植物或动物形成寄生或共生关系(图1)。

图1.主要真菌谱系多样性的代表种[2]。(A)冠珊瑚伞菌Artomyces pyxidata。(B)黄金银耳Tremella mesenterica。(C)花盆阳伞菌Leucocoprinus birnbaumii。(D)平菇Pleurotus ostreatus。(E)银耳Tremella fuciformis。(F) 云芝Trametes versicolor。(G)酿酒酵母菌Saccharomyces cerevisiae。(H)裂殖酵母Schizosaccharomyces pombe。(I) 毛霉Mucor mucedo。(J)玉米黑穗病菌Ustilago maydis。(K)欧灵豪森曲霉Aspergillus oerlinghausenensis。(L) 捕蝇蕈Amanita muscaria。(M) 蝇虫霉Entomophthora muscae。(N)寄生于壶菌异菌体的异菌罗氏菌Rozella allomycis。(O)单毛菌Monoblepharis macrandra。(P)Coemansia braziliensis。(Q) Piptocephalis repens。(R) Mortierella elongata。(S)根霉菌Rhizopus spp。(T) 指状青霉Penicillium digitatum。
历史上,真菌被纳入植物王国,然而,由于真菌缺乏叶绿素,且具有独特的结构和生理特征(即细胞壁和细胞膜的成分),因此它们已与植物分离。此外,真菌在营养生长和营养摄入的主要模式上明显区别于包括动物在内的所有其他生物。真菌从构成生物体的丝(菌丝)的顶端生长,在吸收到菌丝之前,它们从外部消化有机物。
根据DNA分析,大约在15亿年前,真菌从其他生命中分离出来。球囊菌在约5.7亿年前从“高等真菌”(dikaryans)分支而来[1](图2),真菌的进化一直在进行。
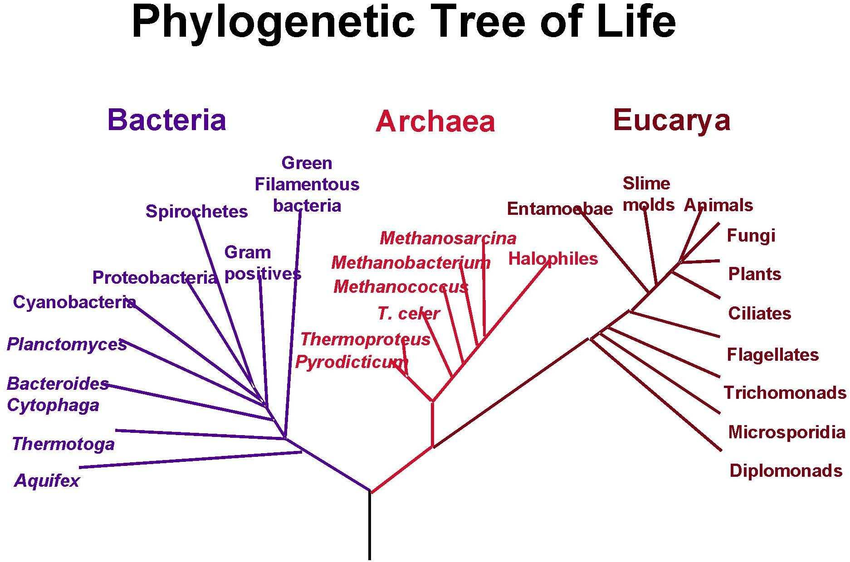
图2.生命之树
这些最早的真菌可能生活在水中,并且有鞭毛最早的陆生真菌化石,或者至少是类真菌化石,大约在6.35亿年前在中国南方被发现(图3)。报道这些化石的研究人员认为,这些类似真菌的生物可能在低温期冰期后的地球大气中扮演了充氧的角色。真菌可能在5亿多年前的寒武纪时期或者6.35亿年前的埃迪卡拉纪就开始在陆地上繁殖,但陆生化石只有在4亿年前的泥盆纪才变得没有争议和普遍[3]。DNA分析的证据表明,所有的真菌都是生活在至少12亿到15亿年前的一个最近的共同祖先的后代。根据化石记录,大约在2.5亿年前,真菌在许多地区变得丰富起来,甚至可能是当时地球上的主要生命形式[1]。

图3. 早期丝状真菌化石。具小泡的玄武岩,岩相薄片。图片来自瑞典自然历史博物馆[7]。
最早具有真菌特征的化石距今约24亿年前,这些多细胞底栖生物具有能够吻合的丝状结构,预计其到达时间约为760-1060 Ma[4]。在古生代(542-251 Ma)的大部分时间里,真菌似乎一直是水生的,由类似于现存壶菌的生物体组成。具有带鞭毛的孢子系统发育分析表明,鞭毛在真菌的进化早期就消失了[5],因此,大多数真菌物种都没有鞭毛。从水生生活方式到陆生生活方式的进化适应需要多样化的生态策略来获取营养,包括寄生、破坏和互惠关系的发展,如菌根和地衣化最近的研究表明子囊菌(Ascomycota)的祖先生态状态为腐生状态[6]。
真菌的早期化石记录很差,因为不像其他生物,真菌结构不容易石化。因此,关于真菌系统发育的理论是以现存真菌的形态特征为基础的。从20世纪初开始,真菌被认为是单系的,所有的真菌都来自于一个失去了光合作用能力的藻类祖先。这就产生了鞭毛真菌,其余的真菌都是从鞭毛真菌进化而来的。鞭毛的丧失和接合菌的进化产生了接合菌门。子囊菌门是由无核的接合孢子产生的。它们之间的联系是一种类似于双足菌的真菌(子囊菌门)。单细胞酵母和复杂的丝状子囊菌成员延长了双核阶段,从它进化而来,被认为类似于现代的外囊菌。人们认为,这种类似外囊菌的祖先是担子菌的祖先[3](图4)。
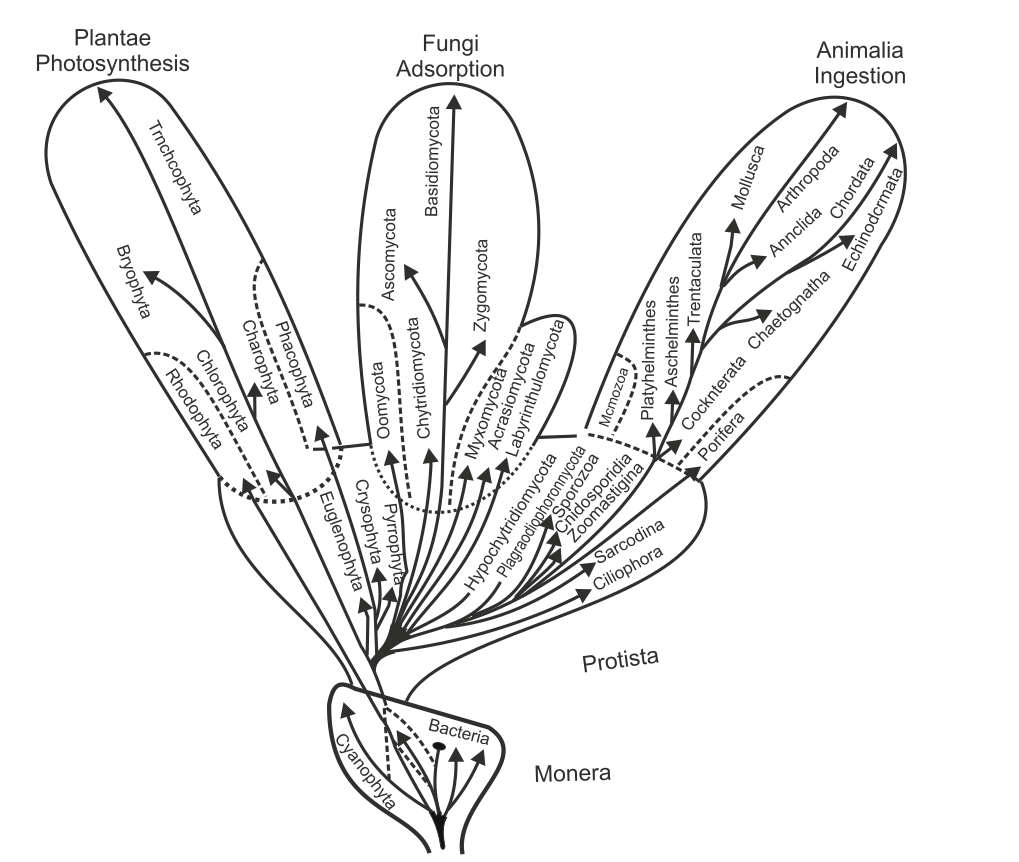
图4. 真菌的起源与分化假说
真菌的单系起源一直到20世纪60年代才被大多数真菌学家接受。但也有真菌学家认为子囊菌属是多系起源,认为红藻是子囊菌属的起源。在20世纪60年代末,卵菌从真菌中分离出来。黏菌也被分离成一个不同的界。真菌分为两个界:鞭毛类真菌全部归入原生生物界(包括壶菌门和黏菌),其余归入菌物界[8]。
随着分子技术、超微结构和生物化学研究的发展,真菌系统发育假说发生了根本性的变化。20世纪80年代末,在这些研究的基础上,壶菌、接合菌、子囊菌和担子菌被划入真菌界。伪真菌中有卵菌门、丝壶菌、网黏菌、破囊壶菌和黏菌。2007年,根据分子系统发育和形态分类学的最新研究结果,提出了一种新的真菌界分类方法,分为1个亚界和7个门[9]。
真菌作为第三个大的分类单元出现,包括那些不属于动物和植物的经典二分法的生物。由于缺乏关于真菌多样性的生物数据,以及整个真菌系统基因组研究相对匮乏,解决真菌树主要类群之间的关系一直是一个挑战[10]。直到基因组学的出现,问题现在才开始得到解决。2021年,科研人员通过综合1500多个公开的基因组数据,向我们提供了一个可靠的系统发育框架来探索真菌进化,并检查冲突的来源和支持真菌系统发育的主干[2](图5)。
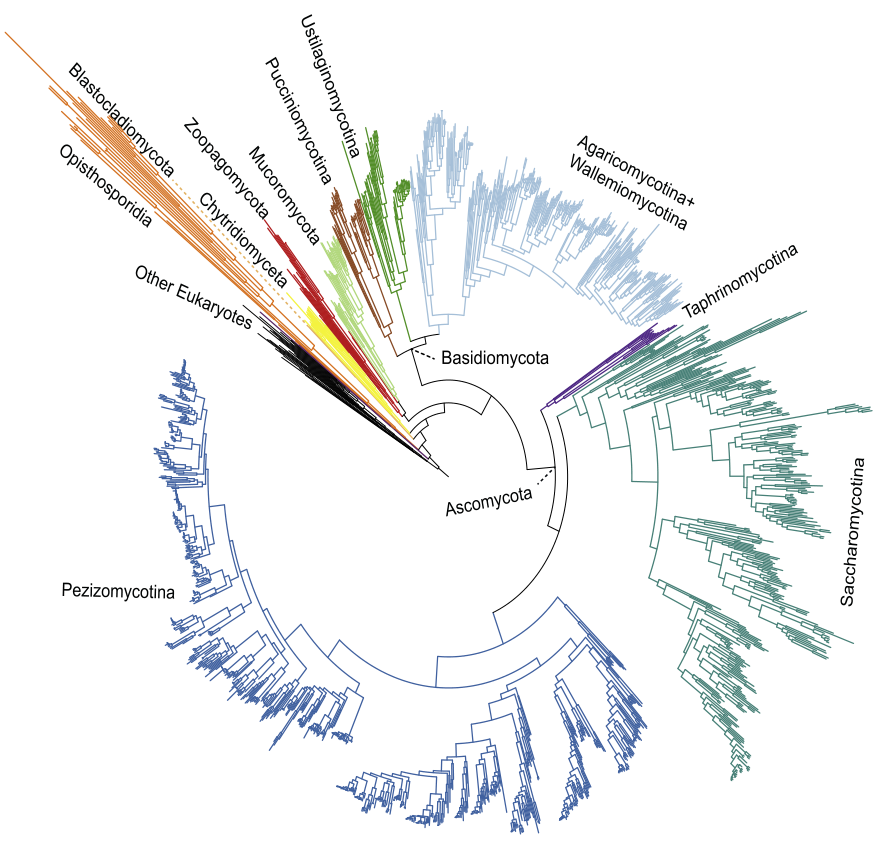
图5. 基于全基因组数据构建的真菌生命之树
基于大量的全基因组信息,真菌系统发育的部分问题都得到了很好的解决,但仍有少数具有挑战性的分支争议没有得到解决。阐明这些尚未解决的关系可能需要大量的数据和分析。尽管最新的基因组级别的系统发育代表了目前支持的最好的假设,但它们总是有可能被修订和改进的。目前尽管有许多类群、基因和分析的研究,但很多关系仍然不明确,这为通过取样额外的分类群和基因来进一步探索有争议的两分区奠定了基础。
真菌虽小,但却与人类的生产生活,生命健康息息相关,与全球的生态系统休戚与共。真菌类群庞大,分布广泛,充满了未知的魅力。值得我们努力一步步去揭开它神秘的面纱。
参考文献
1.Wang, D.Y., S. Kumar, and S.B. Hedges, Divergence time estimates for the early history of animal phyla and the origin of plants, animals and fungi. Proc Biol Sci, 1999. 266(1415): p. 163-71.
2.Li, Y., et al., A genome-scale phylogeny of the kingdom Fungi. Curr Biol, 2021. 31(8): p. 1653-1665.e5.
3.Naranjo-Ortiz, M.A. and T. Gabaldón, Fungal evolution: diversity, taxonomy and phylogeny of the Fungi. Biological Reviews, 2019. 94(6): p. 2101-2137.
4.Bengtson, S., et al., Fungus-like mycelial fossils in 2.4-billion-year-old vesicular basalt. Nat Ecol Evol, 2017. 1(6): p. 141.
5.Lücking, R., et al., Fungi evolved right on track. Mycologia, 2009. 101(6): p. 810-22.
6.James, T.Y., et al., Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature, 2006. 443(7113): p. 818-22.
7.Gan, T., et al., Cryptic terrestrial fungus-like fossils of the early Ediacaran Period. Nat Commun, 2021. 12(1): p. 641.
8.Guarro, J., GenéJ, and A.M. Stchigel, Developments in fungal taxonomy. Clin Microbiol Rev, 1999. 12(3): p. 454-500.
9.Moraes-Barros, N.d. and J.S. Morgante, A simple protocol for the extraction and sequence analysis of DNA from study skin of museum collections. Genetics and Molecular Biology, 2007. 30: p. 1181-1185.
10.Choi, J. and S.H. Kim, A genome Tree of Life for the Fungi kingdom. Proc Natl Acad Sci U S A, 2017. 114(35): p. 9391-9396.








