帝国崛起----蚂蚁社会的形成
引言
谈到蚂蚁,大部分人脑海中出现的场景可能都是孩童时期的夏日午后,趴在树下或路边观察辛勤搬运物资的蚂蚁大军。它们是如此的微小,很容易就会被我们忽视,但它们的数量又是如此的庞大,以至于在任何我们人类出现的地方,几乎都可以见到蚂蚁的存在。当一只蚂蚁落单时,它显得慌乱而笨拙,但当它返回自己的集体,立刻又回归到井然有序的状态。正是这种种的反差,深深地吸引了我们。

图1. 切叶蚁工蚁集体搬运树叶返回巢穴
(图源:https://microbelog.files.wordpress.com/)
蚂蚁属于昆虫纲膜翅目的蚁科,目前包含14172个已命名物种(antcat.org),只占昆虫物种数的13%,但它们的生物量却超出了所有野生鸟类和哺乳动物的总和,约等于地球上全部人类的20% (Schultheiss et al. 2022)。形单影只的蚂蚁简单而又渺小,但当它们聚集在一起时,文明社会的氛围便会显现:一些蚂蚁,例如黑毛蚁(Lasius niger),可以通过放牧蚜虫来让它们产生蜜露供自己食用;行军蚁通过成千上万只工蚁的集体行动来猎杀那些体型远大于自己的小型脊椎动物;收获蚁收集种子并存放在相应的巢室,来帮助自己的家族度过寒冷的冬天;织叶蚁(Oceophylla),通过团队协作将叶片黏连在一起,筑成自己的巢穴;切叶蚁(Atta,Acromyrmex和Amoimyrmex),发展出了先进的农业帝国,通过培植真菌来为自己提供营养。这群起源于1.4亿年前白垩纪的小精灵,到底是如何跨越漫长的演化历史,发展出如此多彩的社会类型,并与人类的文明社会不期而遇?
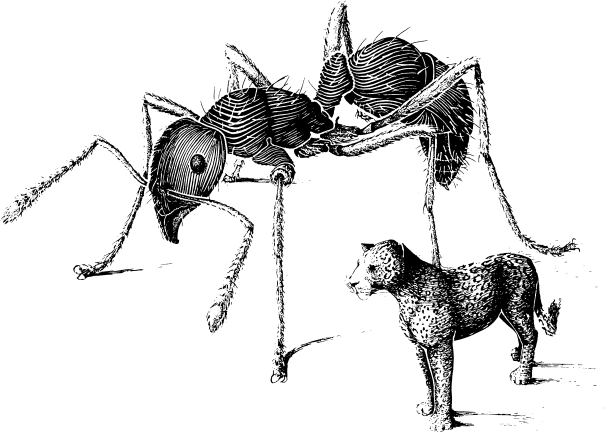
图2. 巴西亚马逊热带雨林中所有蚂蚁和美洲虎的相对质量比
(@Catherine Brown-Wing)
 图3. 蚂蚁社会和人类文明的相似性
图3. 蚂蚁社会和人类文明的相似性
(图源:卫报、维基百科和世界历史百科全书©Simeon Netchev)
01
初识蚂蚁社会--WHAT
自然界中存在着很多种群居的生物,成群结队的大象,集结飞行的鸟群等,但为什么只有少数物种可以被称为社会性动物?广义的社会性动物指的是同一物种按照合作的方式组织起来的群体,其最基本的指标是相互合作和信息交流。以蚂蚁为代表的具有高组织度的昆虫传统上被称为真社会性(eusociality),它们通常具有以下三个特点:第一是世代重叠,即在同一个巢穴中,两代或更多代的成体共存;第二是生殖分工(reproductive division of labor),群体中的一些个体的生殖能力会出现退化甚至丧失,特化出来专门从事劳动的品级(本文出现的品级指的是具有不同形态结构的雌性个体);第三是合作育幼,一些不具有生殖能力的非生殖个体也会参与对群体中幼体的照顾 (Oster and Wilson 1978)。但是从演化生物学的视角出发,真社会性的定义还存在一些模棱两可的地方,比如:对于世代重叠来说,当一些个体因失去其生殖能力而丧失其独立性的时候,是否还能被称为独立的一代;当产生了终身的劳动分工后,有生殖能力的个体专职繁育后代,无生殖能力的个体负责对幼虫的照顾,原本的合作育幼定义还是否适用?真社会性定义中的世代重叠和合作育幼在演化上的意义远小于失去独立性和对更高层次组织的适应性 (Boomsma and Gawne 2018)。

表1. 生命演化过程中的八次重要转变事件
(© 2007 Cold Spring Harbor Laboratory Press)
在20世纪90年代,两位生物学家,史密斯(John Maynard Smith)和绍特马里(Eörs Szathmáry)提出了重大演化转折事件(major evolutionary transitions)。他们提出在生命演化历史上,曾经出现过8次重大的转折性事件,包括从复制子(replicator)到细胞的形成、原核单细胞生物到真核多细胞生物以及独居到社会性等,每一次转变都促进了个体复杂度的提升,将生物带到了更高的生命组织形式:原本独立复制的单元变成了更大组织的一部分,产生了新的劳动分工以及信息传递方式 (Smith and Szathmáry 1997)。从重大演化转折的角度出发,对蚂蚁社会更形象的说法则是超有机体(superorganism),这个概念来源于美国科学家惠勒(William Morton Wheeler),他将蚁巢类比作有机体:蚁后就像生殖细胞(germ cells),负责传递遗传物质,工蚁则如同体细胞(somatic cells),维持机体(蚁巢)的新陈代谢;来自共同祖先的她们通过相互合作来维持机体(蚁巢)的发展;不同品级的分化就如同有机体不同细胞系的分化(图4)。超有机体的概念不仅为蚂蚁社会提供了一个生动的理解视角,而且为研究重大演化转折提供了一个很好的理论框架。相比于作为多细胞生物体基本单元的细胞,作为超有机体基本单元的蚂蚁个体更容易被解构和操作。那么,蚂蚁个体的发育过程,是否和多细胞生物体的细胞分化过程存在相似性?本课题组通过发育期单个体转录组测序的手段对两种蚂蚁(法老小家蚁Monomorium pharaonis和顶切叶蚁Acromyrmex echinatior)进行了探究,随着发育的进行,蚁后和工蚁的转录组特征逐渐开始分化。从而证明了两种不同层次的生命组织形式在发育层面原则的相似性 (Qiu et al.2022)。[“我即是虫群”?以“超个体”的视角看待蚁群]
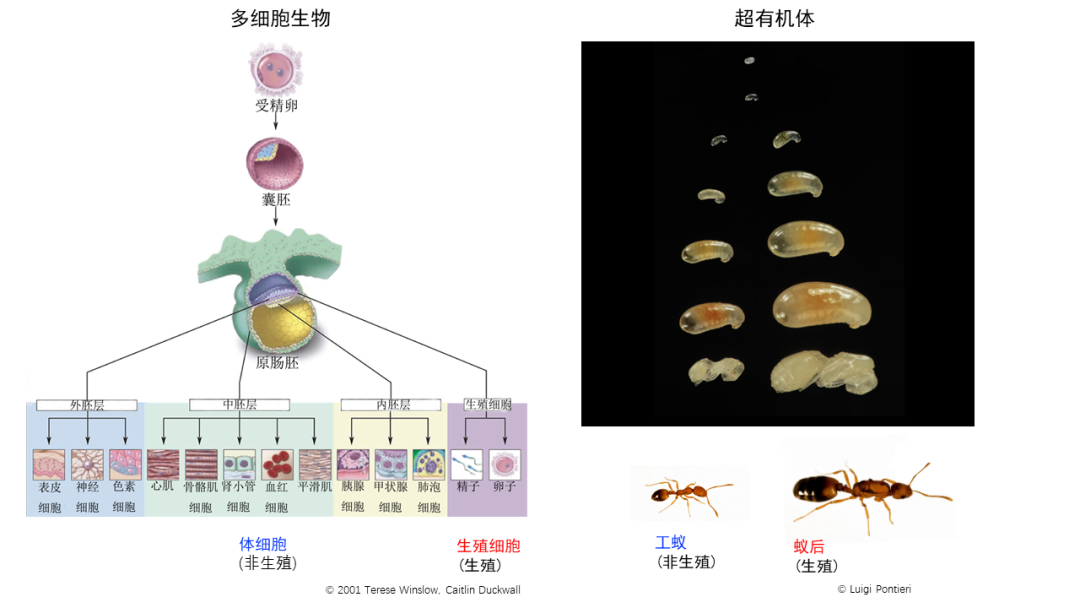
图4. 两种不同生命组织形式的相似性
(©Terese Winslow, Caltlin Duckwall, ©Luigi Pontieri)
02
了解蚂蚁社会的成员--WHO
一个典型的蚂蚁巢穴通常由雌性生殖蚁(交配后便是我们所熟知的蚁后),雌性非生殖蚁(工蚁)和雄蚁组成。雌性生殖蚁通常个体较大,处女生殖蚁(Gyne)通常有两对翅、三个背单眼和发达的卵巢,她们在离开巢穴完成交配后(Queen),便脱去自己的翅,建立属于自己的王国,从此身居王宫深处,一心只为繁殖后代。工蚁(Worker)个体通常小于生殖蚁,她们的翅和单眼退化,卵巢不发达甚至完全丧失,无法生育后代的她们会担负起巢穴内其他的工作。雄蚁(Male)通常只在繁殖季节出现,他们存在的目的就是参与交配,交配完很快就会死掉,他们通常不参与社会分工。与所有现生蚂蚁类群相对应,存在多个已灭绝的蚂蚁物种,保存了这些灭绝物种的琥珀里发现,它们已经存在有翅蚁后和无翅工蚁的分化,说明现生蚂蚁的最近共同祖先很可能也至少拥有这两个品级 (Barden and Grimaldi 2016)。

图5. 一个典型的蚂蚁巢穴,包含蚁后、工蚁和雄蚁(©高琼华)
然而,根据社会的复杂程度,不同蚂蚁也演化出了不同的品级结构,包括工蚁、兵蚁、拟工蚁和蚁后 (Molet, Wheeler, and Peeters 2012)。通常,随着社会向更复杂的方向发展,工蚁会产生更多的亚品级,劳动角色的区分也会更加明确 (Holldobler and Wilson 2008)。但是,品级的发育也会受到很多其他因素的影响,在蚂蚁类群中出现趋同演化等,例如兵蚁的多次独立起源。目前对于品级发育和演化的机制主要有两种观点:一种被称为沙漏模型(hourglass model)(Trible and Kronauer 2021),即品级相关的特征是由体型大小决定的,外界的环境条件比如营养等会影响体型大小,进而影响品级相关特征,从而影响品级分化。另外一种则是二分发育模型(Bifurcating developmental model) (Abouheif 2021),该模型认为不同的品级都来源于完全独立的发育程序(图6)。在大头蚁属(Pheidole)中,胚胎阶段的保幼激素含量决定了是否发育为雌性生殖蚁,而幼虫阶段保幼激素的含量决定发育为工蚁还是兵蚁,不同品级的形成不完全由体型决定。品级发育和演化涉及发育可塑性、发育限制、基因与环境互作和表观遗传调控等学科,随着研究的深入,会对其机制有更深入的认识。
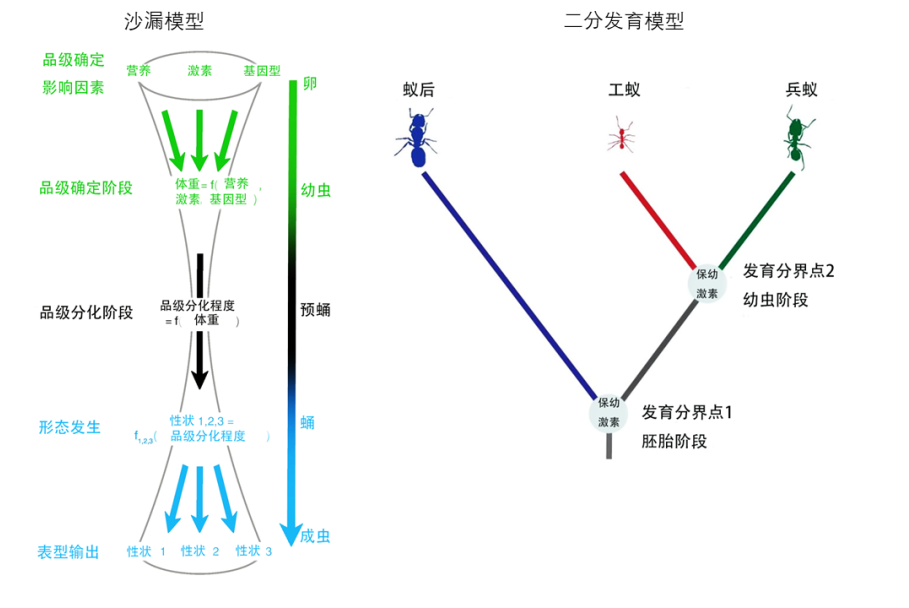
图6. 蚂蚁等级分化的两种模型
(图源:Trible and Kronauer, 2021, Trends Ecol Evol, Abouheif, 2021, Trends Ecol Evol)
03
探究蚂蚁社会的起源--HOW
蚂蚁社会性的起源存在两种假说,一种是准社会性模型(parasocial model),指的是同一代的繁殖个体聚集在一起合作建巢,之后一些个体的行为会偏向繁殖,另一些个体偏向其他任务,最终形成了社会分工。这个假说是基于对一些具有原始社会性胡蜂(Polistinae + Vespinae)的调查,这些物种大部分没有品级分化,通常是由多个雌性共享一个巢穴 (West-Eberhard 1978)。但最近的系统发育基因组学分析指出这些蜂类并不是祖先状态 (Piekarski et al. 2018)。另一种是亚社会性模型(subsocial model),指的是建立巢穴的生殖蚁会延长她们育幼的时间,直到一些后代变成成体并留在巢穴协助育幼。这个假说的一个重要前提是巢穴的单蚁后制。因为大部分蚁后终身只交配一次,这就最大程度保证了产生的后代的亲缘关系,虽然也有一些物种的蚁后会多次交配,但现有研究证明了蚁后单次交配才是她们的祖先状态,因此蚁后多次交配物种的存在不会对亚社会性模型假说产生困扰 (Boomsma 2009)。目前,亚社会性模型也更被学界所接受。
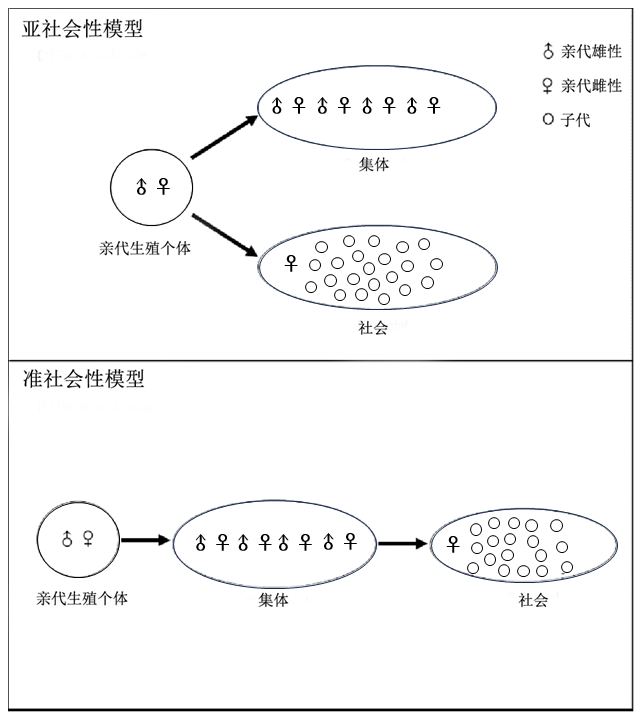
图7. 社会性起源的两种模型
(Chak and Rubenstein, 2019, Curr opin insect sci)
04
社会性形成机制的一些假说--WHY
工蚁的利他行为是如何演化出来的,她们为什么愿意放弃自己的生殖能力,甚至可以为了群体牺牲自己的生命?达尔文(Charles Darwin)在他的著作《物种起源》中曾将工蚁的产生比作“特殊的难题,一开始看起来似乎无法克服,实际上对我的理论是致命的”。他的自然选择理论描述群体中的特征通过变异、筛选和适应性积累来推动物种的演化。但蚂蚁中的工蚁不能生育后代,她们是如何演化的呢,又怎么能分化出不同的亚品级?为此,达尔文提出了家系选择,即自然选择的对象不仅仅是个体层面,对于蚂蚁这类社会性昆虫,自然选择的对象通常是整个群体 (Darwin 1859)。
随着孟德尔遗传定律的发现,英国演化学家汉密尔顿(William Hamilton)于1963年提出了广义适合度理论(inclusive fitness theory) (Hamilton 1963)。他认为自然选择不仅仅影响个体的适合度,还影响与其相关有亲缘关系个体的生存和繁殖成功。这一理论可以用简单的数学公式描述为:rb>c。这里,r代表利他主义者和被帮助者亲缘关系的系数(relatedness),b代表利他主义者的收益(benefit),c代表利他主义者付出的代价(cost)。具体来说,当r×b大于c时,个体倾向于表现出利他行为。这是因为通过帮助亲属间接地提高了其生存概率和繁殖成功率,从而增加了自己的基因在群体中遗传的机率。这一理论很好的解释了社会性动物中利他行为的产生。这一基于亲缘关系的理论,随后也被称为亲选择(kin selection)理论,逐渐成为社会性演化领域被广泛认可的一个底层逻辑,当我们从基因的角度来考虑自然选择时,它为我们提供了合理的推断。但许多研究者也强调,在关注遗传因素的同时不应该忽视生态学参数。
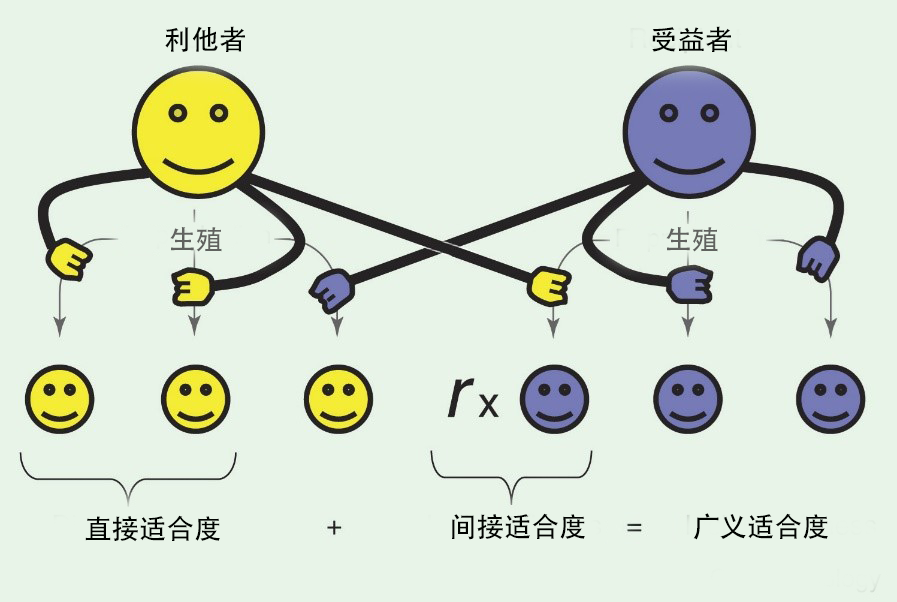
图8. 广义适合度包含直接适合度和间接适合度
(West, Griffin, Gardner, 2007, Curr Bio)
启示
蚂蚁社会诞生至今已有一亿多年,经历了多次环境的剧变,但它们成功应对了环境的选择压力,现在已经发展成了最繁盛的生物类群之一。在蚂蚁内部,也演化出了迥然不同的社会结构。因此,研究蚂蚁社会形成和演化,可以帮助我们更好的探究生物演化的机制、规律和影响因素。蚁后和工蚁具有完全相同的基因组,但是却拥有完全迥异的形态、行为和生理特征(更长的寿命),是表观遗传学极佳的研究对象。蚁后完全不遵守生殖力-寿命权衡(reproduction-lifespan trade-off)的现象,也为长寿相关的研究提供了独特的视角。超有机体概念的提出,不仅使得我们更好地理解蚂蚁的社会性,更是为建立不同生命层次的相关性以及发掘原理上的相似性提供基础,帮助我们理解多细胞生物中细胞的互补整合与协同。
参考文献
Abouheif, Ehab. 2021. “Ant Caste Evo-Devo: It’s Not All about Size.” Trends in Ecology & Evolution 36 (8): 668–70. https://doi.org/10.1016/j.tree.2021.04.002.
Barden, Phillip, and David A. Grimaldi. 2016. “Adaptive Radiation in Socially Advanced Stem-Group Ants from the Cretaceous.” Current Biology 26 (4): 515–21.
Boomsma, Jacobus J. 2009. “Lifetime Monogamy and the Evolution of Eusociality.” Philosophical Transactions of the Royal Society B: Biological Sciences 364 (1533): 3191–3207. Boomsma, Jacobus J., and Richard Gawne. 2018. “Superorganismality and Caste Differentiation as Points of No Return: How the Major Evolutionary Transitions Were Lost in Translation.” Biological Reviews 93 (1): 28–54.
Darwin, Charles. 1859. On the Origin of Species. On the Origin of Species. Oxford University Press.
Hamilton, W. D. 1963. “The Evolution of Altruistic Behavior.” The American Naturalist 97 (896): 354–56.
Holldobler, Bert, and Edward O. Wilson. 2008. The Superorganism: The Beauty Elegance And Strangeness Of Insect Societies. WW Norton.
Molet, Mathieu, Diana E. Wheeler, and Christian Peeters. 2012. “Evolution of Novel Mosaic Castes in Ants: Modularity, Phenotypic Plasticity, and Colonial Buffering.” The American Naturalist 180 (3): 328–41.
Oster, George F., and Edward O. Wilson. 1978. Caste and Ecology in the Social Insects. Princeton University Press.
Piekarski, Patrick K, James M Carpenter, Alan R Lemmon, Emily Moriarty Lemmon, and Barbara J Sharanowski. 2018. “Phylogenomic Evidence Overturns Current Conceptions of Social Evolution in Wasps (Vespidae).” Edited by Keith Crandall. Molecular Biology and Evolution 35 (9): 2097–2109.
Qiu, Bitao, Xueqin Dai, Panyi Li, Rasmus Stenbak Larsen, Ruyan Li, Alivia Lee Price, Guo Ding, et al. 2022. “Canalized Gene Expression during Development Mediates Caste Differentiation in Ants.” Nature Ecology & Evolution 6 (11): 1753–65.
Schultheiss, Patrick, Sabine S. Nooten, Runxi Wang, Mark K. L. Wong, François Brassard, and Benoit Guénard. 2022. “The Abundance, Biomass, and Distribution of Ants on Earth.” Proceedings of the National Academy of Sciences 119 (40): e2201550119.
Smith, John Maynard, and Eörs Szathmáry. 1997. The Major Transitions in Evolution. Oxford: Oxford Univ. Press.
Trible, Waring, and Daniel J.C. Kronauer. 2021. “Hourglass Model for Developmental Evolution of Ant Castes.” Trends in Ecology & Evolution 36 (2): 100–103.
West-Eberhard, Jane. 1978. “Polygyny and the Evolution of Social Behavior in Wasps.” Journal of the Kansas Entomological Society, 832--856.








