Science专刊论文解读:全基因组重测序揭示了狒狒的由雄性驱动的网状演化历史
狒狒是人们比较熟悉的灵长类动物,特别是在很多与非洲动物有关的的纪录片中,它们让人印象深刻。来自奥胡斯大学、贝勒医学院和莱布尼茨灵长类动物研究所等单位的研究人员对19个代表性地区的225只野生狒狒进行了高通量基因组测序,研究了它们的遗传结构模式,并有一些重要的发现。该工作作为灵长类基因组计划的扩展项目,成果已经以“ Genome-wide coancestry reveals details of ancient and recent male-driven reticulation in baboons”(全基因组共祖分析揭示了狒狒古代和近期由雄性驱动的遗传混合)为标题的论文在著名的学术期刊《科学》(Science)上发表。
1. 狒狒和它们的社会制度
狒狒都有长长的,像狗一样的吻(鼻子和嘴),尖锐的牙齿、厚厚的毛发、以及标志性的红屁股。生活在草原、悬崖和戈壁上,以各种各样的植物性和动物性的食物为生。它们是狒狒属(Papio)动物,也是旧世界猴科的23个属之一,一共包含六个物种,分别是几内亚狒狒(P. Papio)、大狒狒(P. ursinus)、东非狒狒(P. anubis)、阿拉伯狒狒(P. hamadryas)、黄狒狒(P. cynocephalus)和金达狒狒(P. kindae)。绝大多数狒狒都生活在非洲,仅有部分阿拉伯狒狒生活在阿拉伯半岛西南部。不同物种的狒狒体型有一定差异,最小的金达狒狒体长仅50厘米,体重仅14公斤。而最大的大狒狒体长可达120厘米,体重40公斤。不同物种的毛发颜色和其他局部特征也有一些差异。之前的研究已经在狒狒基因组内识别了很多不同来源的杂交区域。同时,基于核DNA、线粒体DNA和表型的系统发育研究结果之间具有很大差异。这说明狒狒属物种经历了非常复杂的种间杂交和“网状”演化过程。人类和其它猴子也被认为曾经过类似的演化过程,因此研究狒狒的演化过程也是我们认识自身演化历史的重要方式。然而,对于它们的演化历史和种间杂交机制目前尚缺乏系统的研究。
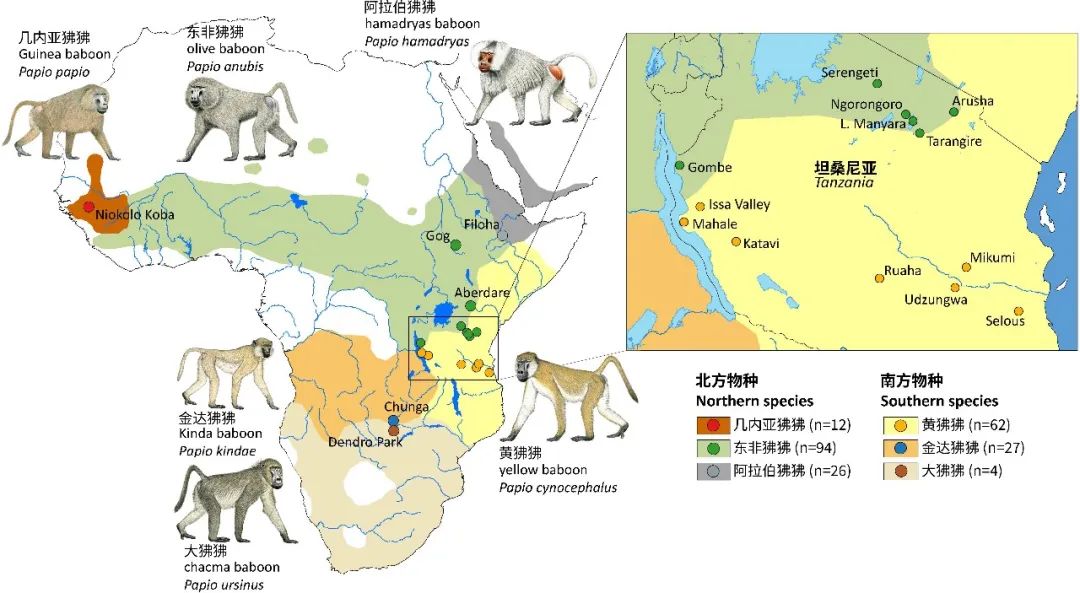
六种狒狒的样貌和分布区域(原图作者:Erik F. Sørensen等)
2. 杂交的狒狒
狒狒复杂的种间杂交和“网状”演化历史是源于它们独特的社会结构和复杂的社会性行为。为了研究狒狒的演化历史,首先要了解狒狒的社会制度。
狒狒具有等级森严的社会制度。科学家们甚至使用“部队”一词形容纪律严明的狒狒群落。大多数狒狒部队都由一个雌性狒狒领导,遵循严格的母权制度。当一只雌性狒狒成年时,通常会立即获得常仅次于她母亲的地位,这种“军衔“的继承相当稳定,往往可以跨越许多世代。未成年的雌性地位总是低于成年雌性,但是未成年雌性之间的地位也是由母亲的地位高低决定的。
有趣的是,部队中雄性的地位争夺是完全不同的模式。未成年雄性的地位是靠着他们的年龄、体型和战斗力争取的,母亲的地位对于雄性之间的竞争几乎没有什么作用。成年雄性的地位却决于他们拥有多少后代,后代越多则他们的地位越高。同时地位越高的雄性往往能更加容易地与雌性交配从而留下更多地后代。
狒狒部队的等级和规矩虽然森严,但也不是一成不变的。相当多的雄性狒狒和部分雌性狒狒会认为自己值得更高的社会地位。雌性狒狒通过主动付出更多的劳动来获得其它成员的认可,这种方式虽然效率很低,但好在安全。而年轻的雄性则可能结成联盟,共同“起义”,向高等级的雄性发起挑战。起义成功的奖励是丰厚的,胜利者往往能直接取代失败者之前的地位。但高收益也伴随着风险,失败者可能会拖着受伤的身体被驱逐出“部队”。这些流浪的狒狒往往还是不停尝试融入其它部队。他们会向其它部队中的雄性发起挑战,如果成功的话同样可以取代失败者在新部队中的地位。实际上雌性狒狒也偶尔会出现跨部队的迁移和扩散,不过这种现象产生的原因目前仍不十分清楚。
研究人员曾在不同种狒狒栖息地的重叠区域发现了大量的自然杂交个体。它们往往具有更大的体型,体现出了杂种优势。这些杂交个体很可能是流浪狒狒融入其它物种“部队”或者雌雄流浪狒狒在野外“邂逅”的产物。
狒狒容易迁移和扩散的特性使得自然杂交时常发生,这很有效地抵抗了物种间地遗传分化。同时,狒狒雌雄之间不同的迁移和扩散模式给分析狒狒的演化和杂交历史造成了很大困难,基于母系遗传(线粒体DNA)、父系遗传(Y染色体DNA)和不区分母系和父系(常染色体)的演化历史分析可能会得到完全不同的结果。因此回溯真实的演化历史必须要综合考虑基于各种数据得到的结果。
3. 基因组揭示遗传信息
研究人员对19个代表性地区的225只野生狒狒进行了高通量基因组测序,来研究狒狒属物种的的群体遗传学和跨物种遗传交流。
研究结果显示,六种狒狒首先可以被分成北方狒狒和南方狒狒两个大支脉。北方狒狒包括阿拉伯狒狒、东非狒狒和几内亚狒狒三个物种;南方狒狒包括金达狒狒、黄狒狒和大狒狒。遗传结构分析则显示了东非狒狒和黄狒狒具有比较复杂的遗传背景和演化历史。其中,来自西部Mahale和Katavi的黄狒狒比东部Mikumi的狒狒体型小一些,有些个体研究周围还有粉红色的皮肤,这些都跟金达狒狒类似。而遗传结构和共同祖先矩阵也显示,西部黄狒狒与金达狒狒的遗传相似性大于它们与东部黄狒狒的相似性。这是跨物种遗传交流存在的有力证据,也是种间杂交的遗留下来的痕迹。研究团队使用遗传变异数据重新构建了不同地理种群的种群规模变化历史。结果显示,从距今10万年至距今100-120万年前,各狒狒种群的规模都是相同的。直到距今10万年,南北方物种开始分化之时,北方物种的群体规模开始明显低于南方种群。这一结果说明,狒狒属物种的祖先发源于非洲南部,然后向北扩散,再扩散至非洲西部。在一系列的扩散事件发生的同时,遗传多样性也因遗传漂变出现下降,导致了新形成的种群规模的下降。现在位于狒狒栖息地最西北的几内亚狒狒,是那些一直在不断扩展栖息地范围的“先锋“们留下的后代。
此前的研究人员曾经认为金达狒狒是北方狒狒和南方狒狒杂交产生的新物种。然而该研究的结果显示金达狒狒具有非常独特遗传结构,与南方狒狒和北方狒狒都不同。基于Y染色体遗传信息构建的演化树认为金达狒狒是所有其它狒狒的姊妹演化枝,而基于常染色体和X染色体遗传信息构建的演化树则认为金达狒狒属于南方狒狒的一种。金达狒狒标志性的Y染色体单倍型在西部黄狒狒中同样存在,而北方狒狒都没有这个单倍型。同时,金达狒狒没有任何北方狒狒的线粒体DNA序列。最后,金达狒狒与不属于狒狒属的狮尾狒共享更多的Alu遗传突变。这些证据都说明金达狒狒不太可能是杂交产生的新物种。综合以上证据,作者们推断:金达狒狒可能保留了更多的祖先遗传物质和古老的外貌形态。为了给这一论断提供证据,作者们进一步分析了这些狒狒的线粒体DNA所携带的遗传信息。化石证据表明狒狒起源于南部非洲。而对线粒体DNA的分析显示,金达狒狒线粒体DNA直系的分布范围正好包含了狒狒的起源地,支持了上述推测。
进一步的研究发现,在东非狒狒和黄狒狒的栖息地交界处,两个物种的种群之间有大量的共同祖先序列,这说明两个物种在该地区持续发生着种间杂交。另外,来自Mahale和Katavi的西部黄狒狒与东部黄狒狒、金达狒狒和东非狒狒之间都存在着大量的共同祖先序列,说明西部黄狒狒种群同时接收了来自两个以上狒狒种群的遗传物质输入,这是该现象在狒狒中首次被发现。
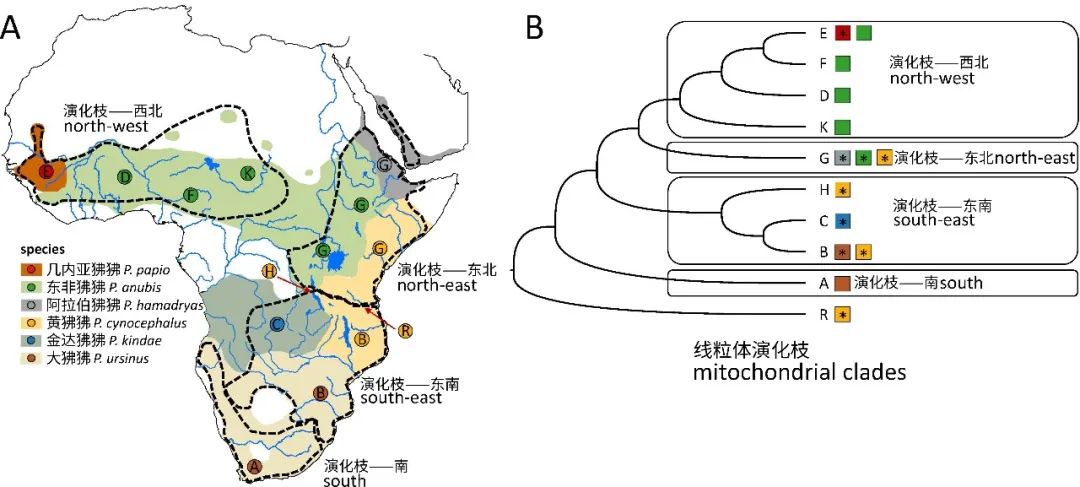
狒狒线粒体DNA支系分布区域和系统发育树(演化树)(原图作者:Erik F. Sørensen等)
4. 更多的信息
该研究还通过比较X染色体和8号染色体(一条与X染色体长度相似的常染色体)的遗传物质来源差异,尝试揭示狒狒中常见的雄性迁移扩散现象对其演化历史的影响。这一方法的原理是雄性只携带一条X染色体,却携带两条(1对)8号染色体,因此,雄性迁移扩散对于8号染色体遗传组成的影响要远远大于对X染色体的影响。
结果显示,最具有说服力的例子出现在金达狒狒和西部黄狒狒之间。西部黄狒狒与金达狒狒在X染色体上的共享祖先序列比在8号染色体上多,而金达狒狒在X染色体上与西部黄狒狒共享的序列更是达到了8号染色体上的两倍。同时,东部黄狒狒与西部黄狒狒共享的X染色体序列比8号染色体多。这些结果共同表明,雄性东部黄狒狒迁移到了金达狒狒的领地,通过杂交将自身携带的遗传物质输入仅东部黄狒狒的基因库中。造成了西部黄狒狒 与东部黄狒狒明显不同的遗传组成。
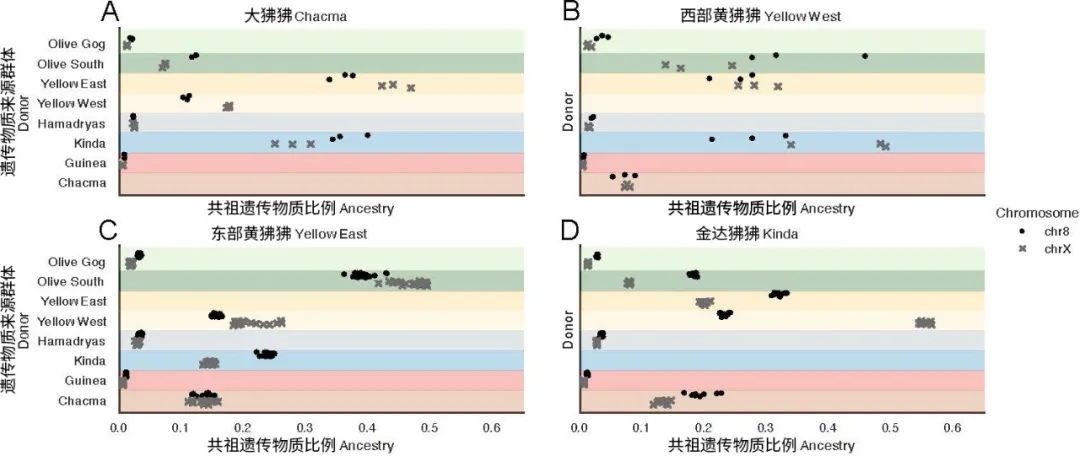
大狒狒、西部黄狒狒、西部黄狒狒和金达狒狒8号染色体和常染色体的遗传组成(原图作者:Erik F. Sørensen等)
最后,该研究还分析了可能与特定环境适应性有关的遗传突变。最吸引人的发现是所有阿拉伯狒狒和几内亚狒狒都拥有一个发生在Neurexin 1(NRXN1)基因上的突变,该突变会改变NRXN1基因的蛋白质编码。而所有其它狒狒都没有这个突变。NRXN1基因与动物的社会行为有关,在小鼠中敲除这个基因会导致明显的雄性攻击性变化。阿拉伯狒狒和几内亚狒狒由于和其它狒狒完全不同的社会组织。阿拉伯狒狒或几内亚狒狒“部队”并不是雌性领导的。他们的部队有许多“小家庭”组成,每个“小家庭”包含一只雄性和几只雌性。在“小家庭中”,雄性狒狒表现得控制欲极强,当雌性走得离它太远时,它会跑过去把雌性抓到身边并“家暴”雌性。同时,雄性之间还经常出现争夺雌性的战斗。该研究关于NRXN1基因突变的解释了雄性阿拉伯狒狒和几内亚狒狒更加好斗的原因。此外,该研究还发现了一些可能分别与金达狒狒独特的毛色特征、体型较小和雌雄差异较低有关的遗传突变。
综上所述,狒狒的遗传结构模式很可能是研究其他灵长类物种群结构的宝贵模型,如非洲绿猴(Chlorocebus)和猕猴(Macaca)。人类祖先的出现、分化和混杂的时间跨度与狒狒的演化历史十分相似,因此该研究也可以作为研究人类演化历史的重要参考。








