NEE论文解读:性染色体跨越八千万年的演化故事
非人灵长类是地球上距离我们最近的亲戚,通过对它们的研究,我们可以更深入地了解人类的起源、演化以及疾病等。尽管它们与人类同属灵长目,但在漫长的演化过程中,这些物种却演化出了各种不同的婚配系统,部分物种还演化出了明显的外观和体型上的两性差异。虽然人类以及部分离人类最近的类人猿的性染色体早已被解析,但目前大部分的灵长类性染色体乃至基因组的信息仍然未知。这些灵长类的性染色体上有什么基因,这些基因又和物种间的差异以及同一物种内雌雄表型差异有什么关系?近日,浙江大学生命演化研究中心张国捷团队,利用全基因组测序数据,重构了灵长类动物性染色体的演化历程。上述相关研究成果发表于《自然-生态与演化》(Nature Ecology & Evolution)。
不一样的X和Y染色体
我们性别差异的遗传学基础来自于性染色体的差异。男性个体拥有在形态、长度等方面都存在巨大差异的X和Y两条性染色体,而女性个体则具有两条X染色体。与常染色体相比,X和Y染色体最大的差别是它们减数分裂形成生殖细胞时只能在染色体端部的假常染色体区域(Pseudoautosomal Region,PAR)处发生联会重组,而其他区域已经在X、Y染色体间发生分化,很难重组(也就是重组抑制),这些区域被称为性别分化区域(Sex Differentiated Region, SDR)。以人为例,人的X染色体长度约为156 Mb,包含有850多个蛋白编码基因;而Y染色体则发生了大量的退化,长度仅剩57 Mb,只有X染色体长度的1/3多一些,只剩下48个蛋白编码基因。
要了解Y染色体为什么会退化,我们首先得知道它是怎么演化来的。目前的主流学说认为性染色体起源于一对祖先常染色体。在演化过程中,Y染色体上出现了性别决定基因,在自然选择的作用下,性别拮抗的突变在性染色体上连锁富集,最终导致与X染色体间的重组抑制。同时,重组抑制也导致了Y染色体无法通过重组将中性或略微有害的突变清除,伴随着重复序列的累积急剧退化。研究表明,人类的Y染色体起源于距今约1.8亿年前真兽亚纲(如人和小鼠)和后兽亚纲(如袋鼠、考拉)的最近共同祖先,且不同的哺乳动物在演化后最终保留下来的Y染色体差异很大。如前文所说到的一样,人的Y染色体只有X的1/3,而跟人最近的黑猩猩的Y只有其X的20%,小鼠则相对更长一些(约为X的一半)。
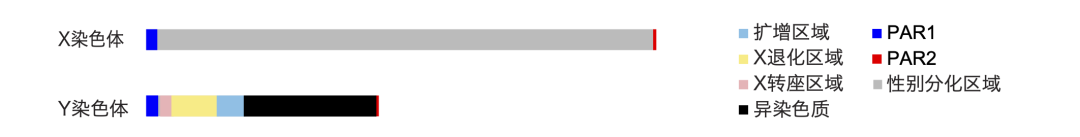
人的X染色体与Y染色体。除两个还能重组的同源区域(PAR1和PAR2)外,Y染色体其他区域为根据相应类别序列所占比例进行汇总标注,不代表真实位置(周旸 绘)
不同物种的Y染色体差别很大
浙江大学生命演化研究中心的张国捷教授和深圳华大生命科学研究院的周旸研究员等利用长读长的测序技术,新构建了19个灵长类物种高质量X和Y性染色体序列,获得了更具有广泛代表性灵长类物种的性染色体信息,完全覆盖了灵长目中的所有的两个亚目(原猴亚目和简鼻亚目),并以树鼩为灵长类的外群,分析时间跨度长达近八千万年。
该研究显示,即使是和人亲缘关系最近的灵长类,在经历了漫长的演化后,不同物种间Y染色体的差异也已经极大。例如,原猴亚目懒猴科中的小懒猴拥有灵长类中最长的Y染色体(约109.66 Mb,约为其X染色体的60%),而最短的是猴科中阿拉伯狒狒的Y染色体(仅约16.24 Mb,为其X染色体的10%)。
研究还发现,除了异染色质区域(主要由卫星序列这种短重复单元构成)外,Y染色体的长度差异主要由扩增区域(Ampliconic region)的大小差异导致的。这类型Y染色体区域主要由大段高度相似的重复序列构成(序列间的一致性高达99%),常形成大型的回文结构(即结构相同、方向相反的序列结构),并包含大量与雄性生殖繁育有关的基因(如DAZ、TSPY基因簇)。值得注意的是,这些扩增序列区域和扩增基因有部分在灵长类的最近共同祖先就已经发生了系统性的扩增,但还有大量的序列是在不同的类群的祖先各自发生扩张,并可能与该类群受到的特定选择有关。例如,DDX3Y这一与精子发育过程有关的Y基因只在狭鼻小目中发生了扩增,这可能与该类群物种多夫多妻的婚配方式导致的激烈的精子竞争选择压力有关。
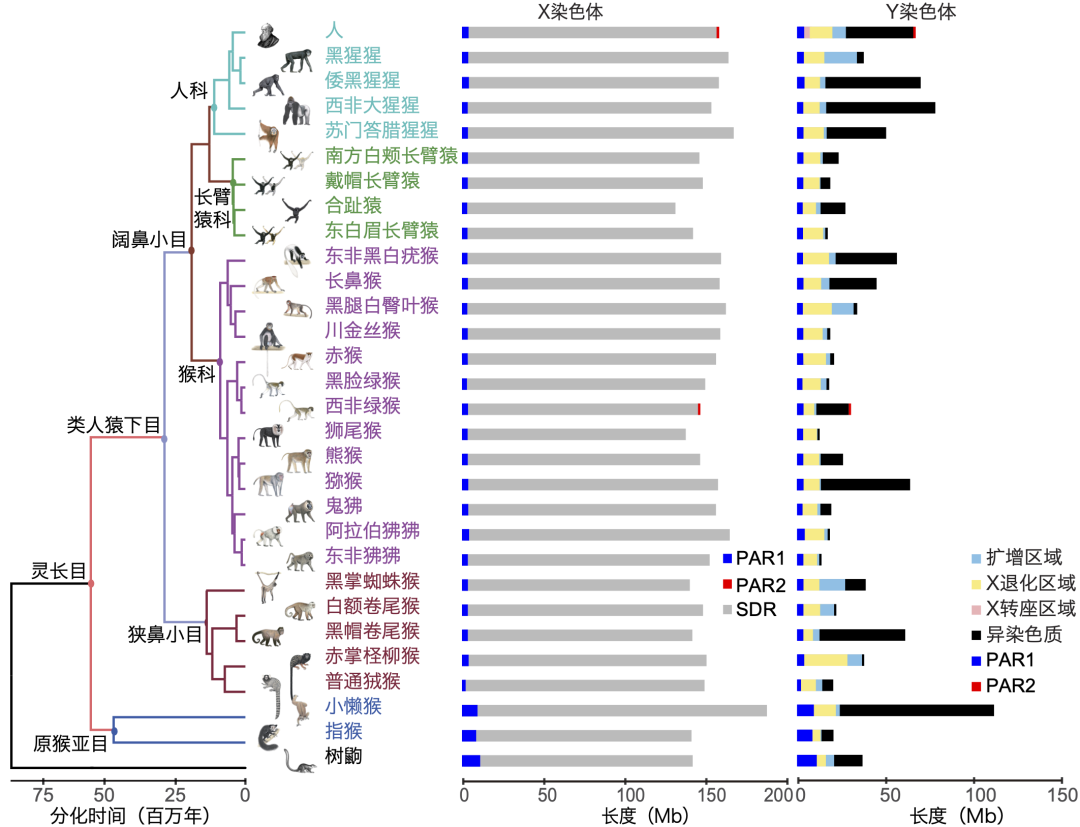
29种灵长类动物的系统发育关系,以及性染色体的长度和组成,灵长类图片由Stephen D. Nash绘制(周旸 等 绘)
雄性更容易自闭可能是性染色体演化导致的副产物
由于重组抑制的作用,灵长类Y染色体上绝大部份的祖先基因都在演化过程中迅速累积了大量的突变,发生了功能性退化甚至丢失,基本上只有十分之一的基因被保留下来,而且不同物种间保留下来的基因也有一定的差别。
然而,一些从祖先常染色体上遗传演化而来的Y基因,由于其功能的重要性在大部分物种中得以被保留下来。这些基因有与物种的生殖发育有关(如性别决定基因SRY),也有参与到生物个体一般的转录翻译调控过程中(如UTY)。
在灵长类中最引人注意的Y基因之一是NLGN4Y——它在29个灵长类中一半的物种(包括人)都被保留了下来,这暗示着它可能对大多数的雄性灵长类都具有重要的作用。该基因主要负责编码I类膜蛋白且可能与神经系统的功能性突触形成有关。值得注意的是,该基因与其X的同源基因NLGN4X基因间存在一个关键的氨基酸编码突变,已有报道发现这一氨基酸差异与自闭症存在雄性发病偏好存在关联。此次研究发现,NLGN4Y上这个氨基酸编码突变早在类人猿下目的祖先就已产生而且一直保留至今,这意味着该突变的形成可能对类人猿下目的雄性发育有重要意义,而该突变的副作用却也引发了雄性自闭症发病偏好性。
性染色体演化有“断层”
X、Y性染色体上的重组抑制的发生往往不是连续渐进式进行的,而通常是通过诸如染色体大片段的反转或易位产生的,这种现象被称为“演化断层”(evolutionary strata)。这些演化断层的存在,如同地理断层一样,在性染色体上记录着自其产生以来发生的各种各样的演化事件。
从1.8亿年到现在,在形成人类的性染色体的演化过程中发生了多次的重组抑制,一共包含5个演化断层,可以记为“S1-S5”。其中,最古老的三个演化断层即S1-S3早在灵长目物种分化之前就已经形成,在所有灵长类物种中都存在;而较为年轻的两个演化层S4和S5则是在灵长类分化后才形成的。
利用更完善的灵长类谱系性染色体数据,研究人员发现在近八千万年的演化过程中,这29个灵长类物种的X/Y染色体间大大小小的分化事件至少发生了6次,其中形成了数个新的灵长目特异性的演化断层(S4-S6)。其中,原猴亚目物种(如前文提到的小懒猴)的性染色体更为原始:它们具有更长的PAR,保留了灵长类共同祖先性染色体的状态,只具有祖先的S1-S3三个演化断层。而在其他灵长目物种的演化过程中,首先在类人猿下目的共同祖先中演化出了一个新的演化层S4。狭鼻小目(如亚洲的金丝猴)和阔鼻小目(如美洲的黑掌蜘蛛猴)则在物种分歧后经历了进一步的Y染色体退化,又各自产生了一个额外的新演化断层S5。最年轻的S6目前只发现存在在美洲的普通狨猴中。
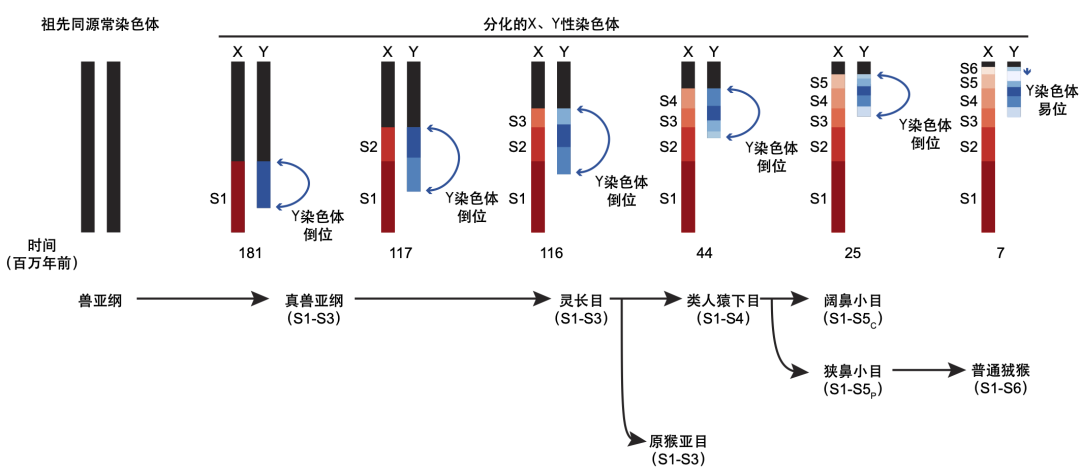
灵长类演化过程中,Y染色体曾发生的演化断层事件(周旸 绘)








