五亿年以后,脊椎动物早已不再是那条文昌鱼
文昌鱼长得像鱼却不是鱼,与包括我们熟悉的硬骨鱼在内的脊椎动物的祖先分道扬镳了5.5亿年。《大话西游》里孙悟空给紫霞仙子许诺了真爱一万年,不过只是这个时间尺度的55000分之一!
脊椎动物在这个我们人类难以想象漫长的时间尺度演化的过程中经历了两次基因组的复制,基因的数目和基因组大小整整比原先多了四倍;随后又经历后续大规模的基因的丢失和基因组序列的重排,从而实现了今天基因的功能,调控方式乃至表型和行为上相比无脊椎动物的高很多的复杂性。这个过程是怎样实现的?一切都要从厦门名产——文昌鱼开始说起…
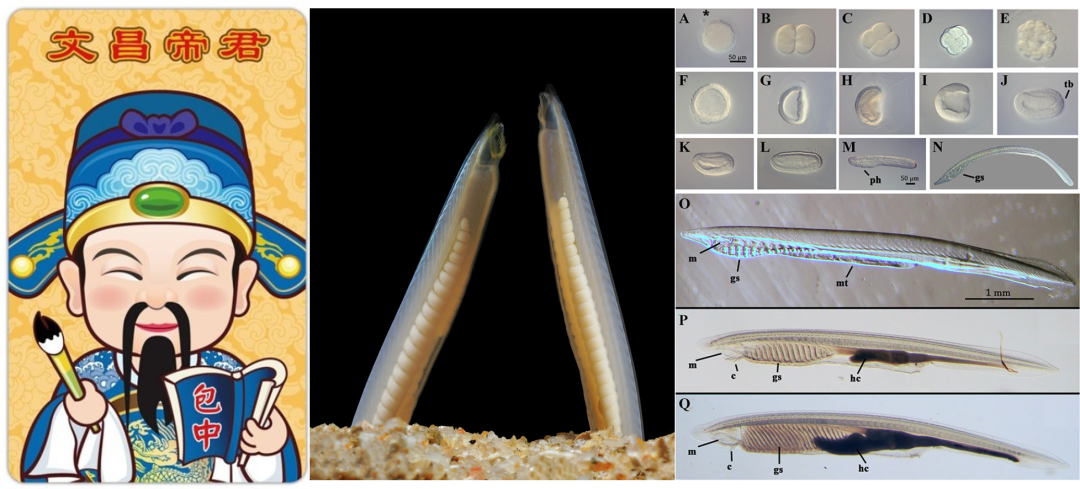
图1 文昌帝和文昌鱼
中图为雌(右)雄(左)佛罗里达文昌鱼右图为文昌鱼从受精卵(A)直至成年(Q)各个发育时期的照片(1)。
从筷子到镊子:
厦门名产在生物学研究中的起起落落
文昌鱼像很多其他我国被研究得很多的动植物一样,原先只是普通老百姓的盘中餐,最开始是被筷子而不是被镊子夹住的。之所以有一个听起来似乎很高大上的名字,是因为古代传说掌管考试的神先文昌君和魁星君的坐骑鳄鱼,在厦门附近死掉以后变成了一座鳄鱼岛,并且尸体上产生了白色的文昌鱼。这名字也与每一年渔民大量捕捞文昌鱼的时间大致是文昌帝诞辰有关。文昌鱼的名字究竟从何而来并不重要,从1774年有第一次正式的科学记录时,关于它到底是一种类似海参或者海胆类的无脊椎软体动物,还是一种鱼,即脊椎动物在很长之间内都饱受争议。1864年, 一位来自俄罗斯的研究生Alexander Kowalesky来到意大利的那不勒斯海边租了一个公寓, 每天在海里捞各种包括地中海文昌鱼在内的海洋动物研究它们的早期胚胎发育。他的发现并未给文昌鱼身份的争议带来任何帮助, 但却是历史上第一次重要的对文昌鱼胚胎发育的详细描述:他发现早期文昌鱼的胚胎类似无脊椎动物海胆,到了幼虫时期又更像是脊椎动物。他的文章在1867年被正式发表以后,立刻迎来了国际关注。这种介于脊椎动物和无脊椎动物的特点提示文昌鱼是一种脊椎动物和无脊椎动物的“过渡”生物,也符合当时德国发育生物学家恩斯特·海克尔(Ernst Haeckel)的胚胎重演论(Haeckel’s doctrine):即演化过程中如果“低等动物”有足够多的时间发育,就能长成“高等动物”。海克尔的这一理论是根据许多脊椎动物的早期胚胎外形极为相似,但一定后期发育阶段开始逐渐形成差异而提出来。所以直至今日,许多人仍然将文昌鱼视作可以理解无脊椎动物向脊椎动物实现飞跃的“活化石”。
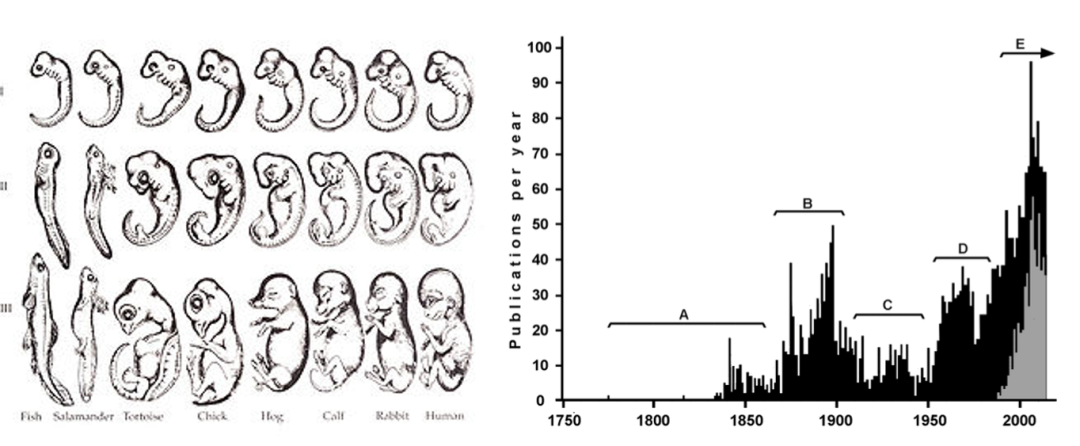
图2 海格尔的胚胎重演论和文昌鱼研究的大起大落
左图为生物学教材中经常提到的恩斯特海格尔的胚胎重演论,他提出脊椎动物不同时期的胚胎形状,提示一个物种的成年状态代表着另一个更“高等”物种的发育过程中的某一个阶段。今天该图的胚胎准确性已受到质疑,理论也早已被推翻;
右图为1774年到2015年间关于文昌鱼研究发表的著作数目随时间经历的起起落落(2)。
然而今天我们回望,不论认为某个现生物种代表了两个类群之间的“过渡”类型,或者认为某些生物类群比较“低等”,脊椎动物或者人类更加“高等”,或是把某个物种认为是“活化石”都是对生物演化过程的误解或是不准确的描述。而海格尔的胚胎重演论更早已被学术界批判抛弃多年,甚至有人质疑其当时胚胎绘图的准确性。”活化石”(living fossils)这个大众再熟悉不过的词,最早被达尔文在《物种起源》中用以描述鸭嘴兽和肺鱼,用以描述现生物种与百万或者上亿年前化石中发现的生物长得很近似的现象,看起来似乎对抗了漫长的时间未发生任何变化。但即使是达尔文当时的原话也是“These anomalous forms may ALMOST be called living fossils”。这一个词ALMOST其实达尔文就在说这些生物类似但并不是所谓的“活化石”。而我们现在也早已知道鸭嘴兽并非是鸟类和哺乳类的“过渡”物种,只是兼具了两者的一些特点,更遑论此“鸭嘴”其实是软组织,和坚硬的鸟喙没有任何关系。著名科普作家Ed Yong 2013年在《科学家》(3)杂志曾经就采访了许多演化生物学家,并以“The Falsity of Living Fossils”(“活化石一词的误用”)指出这个词早就该退休了。因为它很容易给大众造成大众一种误解:就是一些生物可能很长的时间内都不发生演化,并保留了化石一样的形态。然而生物形态上的近似所造成的误解罄竹难书(让我们再想一下鸭嘴兽和鸭子的例子,或者是鲸鱼和硬骨鱼的“鳍”),并且我们今天已经知道只要DNA序列在复制,就会一定几率出错和产生突变,突变和自然选择等其他因素相互作用就在群体水平会不间断地发生演化。而现生的物种类群之间,不论它们表型上看起来有多类似,甚至呈现某种连续的“过渡”,都不可能由一个物种横向“转化”为另外一个物种,否则我们就犯了150多年以前达尔文同一时代的人问达尔文“到底你的祖父还是曾祖父是猴子”的错误了。
当然这些都是题外话,并不能掩盖十九世纪末二十世纪初生物学家们对这种又像脊椎动物,又像无脊椎动物的“鱼”的狂热。尽管对文昌鱼的动物学和胚胎学方面的研究热潮在欧洲的两次世界大战期间被打断,上世纪60-70年代电子显微镜在观察动物组织上的应用,使得对于文昌鱼的形态学研究深入到每一个组织器官,从而研究开始复苏。而从90年代以后基因克隆技术,基因组技术,和遗传操作的发展,使得这个昔日连是不是鱼都搞不清的物种迎来了新的辉煌。
图3 海格尔的插画
尽管海格尔的理论和研究有其历史局限性,但他是一位出色的生物插画艺术家。
童第周、三个荷兰得和佛罗里达海边的披萨
文昌鱼至今不像果蝇,线虫等其他无脊椎动物成为在全球生物学实验室流行的模式生物,主要的原因之一是因为其在实验室养殖条件较为苛刻。这与文昌鱼的生活习性有关。首先,文昌鱼为滤食性生物,主要以悬浮于海水中的单细胞藻类为食物。所以,要实现文昌鱼实验室养殖必须先解决其吃的问题,建立持续稳定的藻类培育体系,而这并不容易。其次,文昌鱼除了在早期胚胎发育阶段和幼体阶段会经历短暂的浮游生活外,后期的生活几乎都在沙中度过,包括吃喝拉撒,所以要确保文昌鱼生活环境质量,必须经常洗沙换水,该过程耗时费力,因而让很多生物学家“知难而退”。而最早解决这一困难, 并且成功实现实验室内文昌鱼饲养繁殖,人工授精的即是我国著名的生物学家童第周。他在上个世纪五十到六十年代山东大学工作期间,利用染色的方法绘制了文昌鱼8细胞和32细胞期胚胎的细胞发育图谱,是世界上早期最重要文昌鱼的发育图谱之一,一直沿用至今。值得一提的是,对于文昌鱼的研究跟对其食用的历史(现在文昌鱼已经是国家二级保护动物,禁止过度捕捞)一样在我国源远流长。西北大学舒德干院士领导的团队1999年,在代表了寒武纪大爆发的著名云南澄江化石群中发现报道类似文昌鱼的“华夏鳗”化石(4),在时间上对其他脊索动物和脊椎动物的祖先的分化年代做了估计。中山大学徐安龙教授和黄盛丰教授利用文昌鱼为模式研究脊椎动物免疫系统的演化(5)和产生了我国白氏文昌鱼的第一张基因组草图(6)。中国海洋大学张士璀教授在文昌鱼和与脊椎动物肝脏和垂体对应器官的研究,尤其他在2020年出版了《文昌鱼演化生物学》一书。最近又由昆明动物所毛炳宇研究员等人绘制的文昌鱼单细胞转录组发育图谱等等(7)。值得一提的是,厦门大学的王义权教授和李光教授团队是世界上最早实现文昌鱼实验室内全年可控繁育(8)、以及文昌鱼遗传操作,即基因编辑敲除和转基因的团队(9)。
而国外近现代对文昌鱼的研究,大部分都与三个“荷兰得”或者他们培养的学生或者博士后息息相关。这三个荷兰得分别是英国牛津大学的Peter Holland, 美国圣地亚哥Scrippt海洋研究所的Linda Holland和Nick Holland。Nick和Linda是一对夫妇,而Peter恰好和他们同姓。Peter在2017年他写的半自传半散文的一篇名为“The dawn of amphioxus molecular biology…”(10)(“文昌鱼分子生物学研究的黎明”)中写道,出于对重要的发育基因Hox 基因的共同兴趣,把这三个未来文昌鱼研究界的三剑客联系到了一起。Nick在得知Peter的兴趣以后,将自己的基金申请完整的寄给了Peter,他们惊奇地发现两人的想法几乎如出一辙。当时Nick摸索出来如何在实验室孵化文昌鱼卵,Linda具有丰富的分子生物学经验,而Peter之前研究和克隆过老鼠的Hox基因序列。在上个世纪90年代,一个远不像今天科研逐步走向平民化,竞争白热化的“春秋”时期,毫无保留地像可能的竞争者共享自己的想法和技术,最终促成他们三人主动合作并走向更宽广的天地: 很快Peter的实验室利用Linda提供的文昌鱼基因组DNA克隆了Hox基因序列,而Linda进一步在Peter的实验室完成了Hox基因的原位杂交。一段很有意思的轶事是,当时Peter(今天他已经是英国皇家科学院院士)把这一结果在瑞士的一次会议中以墙报形式展出,当时Cell杂志的主编,也是创建人之一Ben Lewin经过墙报,看了他一眼,以整个房间都能听到的音量大声问:“What the *** is an amphioxus?”
Peter的这篇发表在正经科研杂志上的趣闻,是我读过的千万篇力求一丝不苟,滴水不漏,信息爆炸的当今科研论文里, 最人性化,最能体会到年轻学者那种十几年来历久弥坚的科研激情的(因此这篇科普这一段写的内容大部分都是他原文的翻译)。这种爷青回的感觉至今读来都可以从屏幕的pdf上扑面而来。Peter他们研究的文昌鱼俗称“佛罗里达文昌鱼”(Branchiostoma floridae),是因为该物种是在美国佛罗里达州的坦帕海湾(Tampa Bay)每年七八月份得以大量捕获。另一个让我印象至今难忘的是,他们的工作需要大量收集不同时期文昌鱼的受精卵,但当时他们并不清楚什么因素促使和具体什么时间文昌鱼的野外种群会产卵。因此他们当时唯一的策略就是“守株待兔”:每天下午三点开始,不顾佛州夏天的温度和湿气,他们的小分队就会带着铲子和筛子在齐腰的海水里挖掘数个小时,然后在傍晚回到实验室,安顿好当天收获的卵,去一家便宜的叫Papa Loui的披萨店用餐。在那家小餐馆里于是诞生了无数的一天体力劳动之后的头脑风暴。我在此处给他们配图复联拍戏杀青以后在土耳其烤肉店的图片觉得再合适不过。

图4 三个荷兰得
图出自Peter Holland的爷青回名作(10): A是当时他们在坦帕湾收集文昌鱼;B是Linda Holland;C是Peter和他的学生以及博后在收集观察文昌鱼卵;D是Peter当时在机场用随身带的实验器材分选文昌鱼卵。
 图5 佛罗里达的披萨店(借复联图复原)
图5 佛罗里达的披萨店(借复联图复原)
基因组和基因编辑时代,文昌鱼走向未来
说了这么多文昌鱼的研究往事,我们说回文昌鱼的分类地位。绝大多数演化生物学的研究都依赖于所研究物种的系统发育关系作为“GPS”,我们也可以俗称“历史定位”。当然这个历史尺度,人类的文明史相比只是沧海一粟。现在我们已经知道,文昌鱼属于头索动物,海鞘等属于尾索动物,它们两者与脊椎动物组成了脊索动物。这里的脊索是指出现在胚胎背侧有弹性的棒状结构: 文昌鱼终生有脊索,海鞘类脊索只存在于早期发育时期,而脊椎动物中,脊索则是脊椎的前体。当然除了脊索,脊索动物其他的特征还包括背神经管,体节咽鳃裂,肛后尾等等。有意思的是,尽管文昌鱼和脊椎动物外形更接近,类似一根透明管子一样外形的海鞘(关于这个物种中国海洋大学的董波教授是研究专家)却系统发育关系与脊椎动物更接近。这一违反直觉甚至让人大跌眼镜的科学发现,即脊椎动物的最近亲缘关系的现生物种长的像个透明管子而不是像条鱼,直到2006年对一种浮游生物尾海鞘(Oikopleura Dioica)的全基因组测定并与其他脊椎动物和文昌鱼的基因序列一同构建产生的系统发育树最终开始完全被学界接受。有意思的是仅三年前在2003年另一个小组测定的另外一种尾索动物玻璃海鞘(Ciona intestinalis)还依然将文昌鱼和脊椎动物的系统发育关系错误地认为更接近。因此2006年发表在《自然》杂志的这项工作标题即为“尾索动物,而不是头索动物是脊椎动物亲缘关系最近的现生物种”(Tunicates and not cephalochordates are the closest living relatives of vertebrates)(11)。
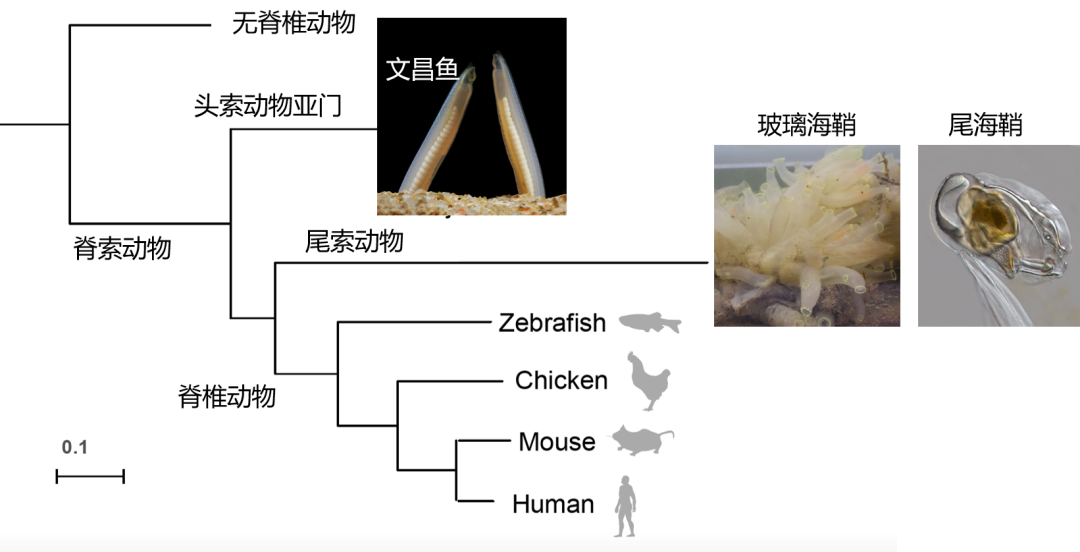
图6 脊索动物系统发育关系
到2008年文昌鱼基因组草图的完成(恰好是发表2003年玻璃海鞘基因组的同一个小组),再次确认了文昌鱼是脊索动物的“基部”物种,即最早与尾索动物和脊椎动物分道扬镳。更重要的是,基因组序列为Susumu Ohno大野乾[11] 早在上个世纪70年代提出的极具前瞻性的假说“脊椎动物祖先经历了两次基因组重复”(the 2R hypothesis)提供了直接的证据。在2018年欧洲文昌鱼(B. lanceolatum, 即文章一开头那个俄罗斯学者在意大利那不勒斯海边研究的那个物种)多组学数据在《自然》杂志上发表。最近我和厦门大学的李光,福建师范大学的黄镇以及西南大学的徐洛浩等联合产生了三种文昌鱼[https://www.pnas.org/doi/10.1073/pnas.2201504120] (中国白氏文昌鱼,日本文昌鱼和佛罗里达文昌鱼),并首次报道了他们的性逆转现象和性别决定系统。这一系列的工作预示着基因组时代的文昌鱼将迎来新的辉煌。
【参考文献】
1. H. Escriva, My Favorite Animal, Amphioxus: Unparalleled for Studying Early Vertebrate Evolution. Bioessays 40, e1800130 (2018).
2. N. D. Holland, L. Z. Holland, The ups and downs of amphioxus biology: a history. Int J Dev Biol 61, 575-583 (2017).
3. E. Yong, The Falsity of Living Fossils. TheSientist (2013).
4. D. G. Shu et al., Lower Cambrian vertebrates from south China. Nature 402, 42-46 (1999).
5. S. Huang et al., Genomic analysis of the immune gene repertoire of amphioxus reveals extraordinary innate complexity and diversity. Genome Res 18, 1112-1126 (2008).
6. Y. Feng, J. Li, A. Xu, "Amphioxus as a Model for Understanding the Evolution of Vertebrates" in Amphioxus Immunity. (2016), 10.1016/b978-0-12-849903-0.00001-4, pp. 1-13.
7. P. Ma et al., Joint profiling of gene expression and chromatin accessibility during amphioxus development at single-cell resolution. Cell Rep 39, 110979 (2022).
8. G. Li, Z. Shu, Y. Wang, Year-round reproduction and induced spawning of Chinese amphioxus, Branchiostoma belcheri, in laboratory. PLoS One 8, e75461 (2013).
9. G. Li et al., Mutagenesis at specific genomic loci of amphioxus Branchiostoma belcheri using TALEN method. J Genet Genomics 41, 215-219 (2014).
10. P. Holland, The dawn of amphioxus molecular biology - a personal perspective. Int J Dev Biol 61, 585-590 (2017).
11. F. Delsuc, H. Brinkmann, D. Chourrout, H. Philippe, Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439, 965-968 (2006).








