种以上分类阶元是否有生物学基础?
提示:快速浏览完这篇文章大概需要10分钟,带着思考大概需要30分钟。这个问题还有许多未解之谜,如果你有兴趣更深入思考,也许值得投入一辈子
前言
在传统的分类法中,分类阶元从物种(Species)、属(Genus)、科(Family),到更大的阶元如目(Order)、纲(Class)、门(Phylum);在我们探讨物种(Species)概念的真实性之后,考虑到物种内部相对缺乏生殖隔离的生物特性,其他的分类阶元可能并不直接具备真实性这一特征,因为它们缺乏遗传物质的自由交换。然而,“种以上的分类阶元是否具有生物学基础?”这一问题也值得我们进行探讨。传统的分类体系为理解生物体的多样性提供了一个很好的组织框架,帮助我们更好地理解和研究生物体之间的关系。在向比“物种(Species) ”更高的分类级别迈进时,如属、科、目等,这些阶元的界定通常涉及更多的形态学、生态学和演化关系。
基于生物的遗传与演化而言,“物种(Species) ”是一个有特殊地位的分类单元。由于物种内部相对无生殖隔离,祖先种群内的广泛杂交使得种内不同个体的系统发生关系总体上呈现出复杂的网状结构;对于不属于同物种的生物体而言,生殖隔离会使它们的系统发生关系基本呈现出复杂的树形结构(在一定程度上也可能会受到跨种杂交、基因渐渗等现象的干扰而呈现零星的网状结构)。因此,对于物种以上的其他分类阶元,我们需要在生命之树(Tree of life)所假定的树形结构的基础来考察它们的划分是否具有生物学基础。
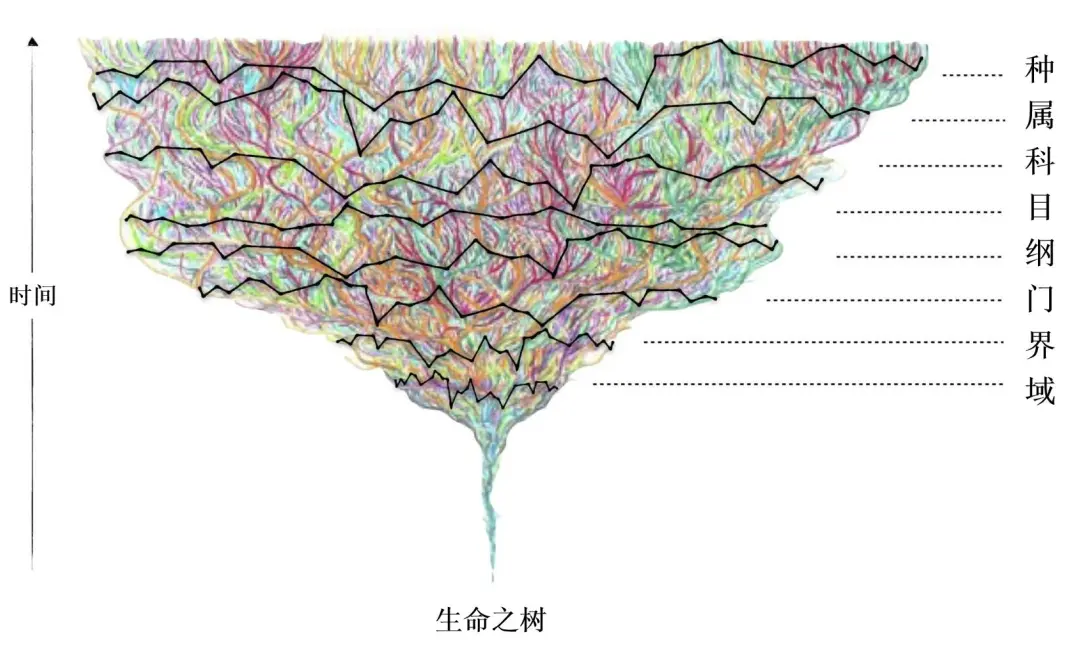
生命之树上的生命体(彩色线条)与传统分类法的分类阶元(黑色线条)的示意图。
(图修改自:Karen Klitz from Swartz and Mishler, 2023.)
01
划分方式的单系性
随着分子生物学技术的发展,特别是基因组学和系统发生学的进步,生命之树(Tree of life)的构建出现了可能。但由于生物体之间的演化关系可能呈现出连续性,而不是清晰的分隔。在某些情况下,物种间的界限可能模糊不清,可能存在相互重叠或渐进过渡的情况,这就使得对不同分类阶元的界定变得更加复杂和模糊。在系统发生树结构下,我们希望每一级的分类阶元都应当代表一个完整的独立分支,即单系群(Monophyly)。大部分的纲(Class)是单系群。例如,我们所熟知的哺乳动物纲(Mammalia)就是一个单系群,它包括了所有哺乳动物的共同祖先及其所有后代。
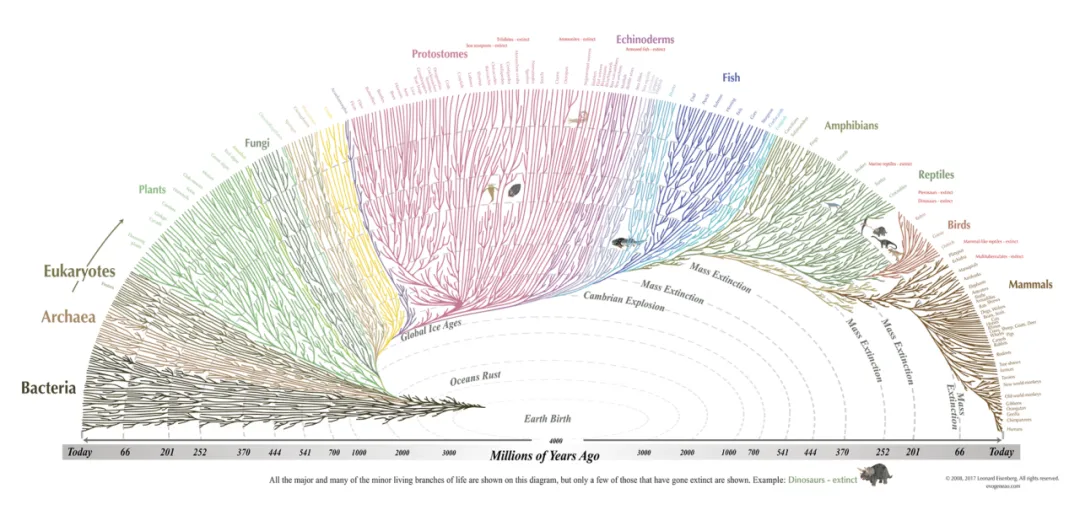
生命之树:不同颜色代表不同类群,哺乳动物(mammals)形成单系群 (图源:evogeneao.com)
但事实上,基于各种形态学、生态学和地理分布等描述性表型特征的传统分类方式不能完全满足形成单系群这一要求。例如,根据过去的传统分类方案,爬行纲被定义为“既非鸟类也非哺乳类的羊膜动物”,因此爬行纲(Reptilia)包含了蜥蜴、蛇、龟鳖和鳄鱼等。然而,爬行纲物种的共同祖先的所有后代还包括所有鸟类和非鸟恐龙,因此传统爬行纲并非单系群;一些分类学者提出了重新定义爬行纲的想法,他们希望将鸟类以及非鸟恐龙包含进来,以形成单系群,或者使用包含非鸟恐龙和鸟类的蜥形纲(Sauropsida)来替代过去爬行纲(Reptilia)的定义。从系统发生关系的角度来看,传统的七阶分类方式并不是很理想,有些类别的命名因其数百年的历史惯性而显得积重难返。
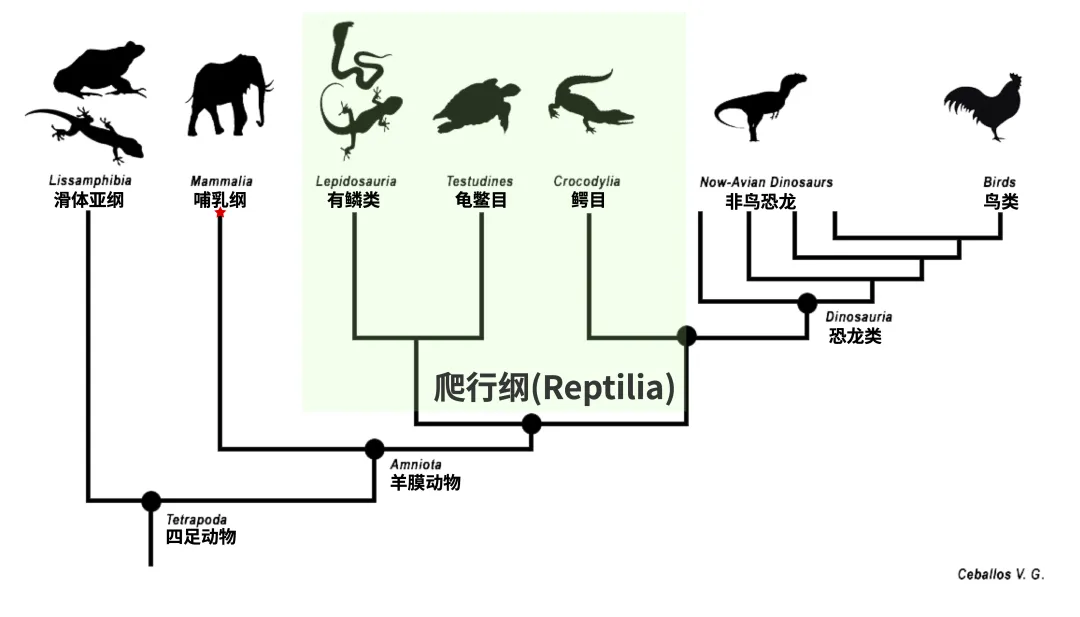
传统分类方案中的爬行纲并非单系群(图源:Ceballos V. G.)
然而,为了有效理清物种多样性的规律和起源,我们又不得不设立各种类别概念对某个或大或小的物种集合加以描述。这时候我们需要在传统的门类单元之间插入更多灵活的划分方式,例如,人们将鸵鸟、鸸鹋等所在的四个目的鸟类定义为平胸总目(Ratitae),因为它们都有扁平的胸骨,无龙骨突,翅膀退化,丧失飞行能力等显著特点。但在最新发布的鸟类系统发生关系中,平胸总目并非一个单系群,分布于拉丁美洲的会飞的䳍形目(Tinamiformes)鸟类与平胸总目鸟类中美洲鸵鸟目(Rheiformes)应当是更为近源物种发生关系。因此,包含了,并非单系群的平胸总目与䳍形目(Tinamiformes)鸟类的古颚下纲(Palaeognathae, 也称古颚总目)应运而生。

古颚下纲鸟类系统发生关系。(Josefin Stiller 和 陈光霁绘)
类似的还有超门(Superphylum)、亚门(subphylum)、小纲(Parvclass)、下目(infra-order)等无数具体概念,但这种细致的类别划分同样要求其内部的所有物种共同构成一个单系群。也就是说,该类别的生物所共享的特征必须继承自它们的共同祖先,而非趋同演化所导致的,这样的特征被定义为同源特征。因此,我们不能因为海豚和蝙蝠都有回声定位的功能而单独地将它们化为一个类别,因为它们的这种生物特征是各自独立演化的结果。
因此,基于演化的分类系统必须反映其产生的自然秩序。也就是说,各种各样的生物特征,从基因序列到复杂表型,它们都要受到系统发生关系的支配和约束,自然选择虽然能改变方方面面的生物性状,但不能给现有的地球生物带来一套全新的遗传密码,或者给现生动物替换出一套截然不同的神经系统。这使得我们充分相信,那些高度保守的基础特征是所持有者继承自它们的共同祖先的结果。如果像脊索、脊椎这样奠定一个门类的主要特征可以独立地多次起源的话,那我们由此定义出的脊索动物门、脊椎动物亚门就毫无演化意义了。
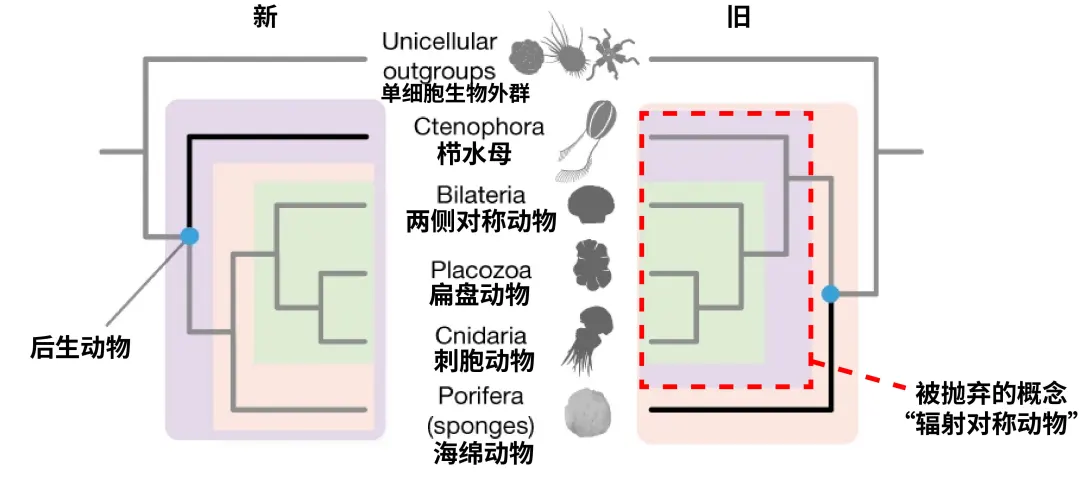
后生动物主要类群的系统发生关系示意图。左图为新的系统发生关系,支持栉水母为其他后生动物的姊妹裙;右图为以往的系统发生关系,依据辐射对称提出的辐射对称动物(已逐渐被抛弃的概念)。(编辑自https://www.nature.com/articles/s41586-023-05936-6的Figure 1)
与此同时,我们也会不断修正一些认知,例如过去我们根据辐射对称的特征将水母等刺胞动物与栉水母生物划分为辐射对称动物,而基于基因共线性的证据显示栉水母应当是其他所有后生动物的姊妹群,因此无法与刺胞动物划为同一门类,于是近年来,“辐射对称动物”这一概念逐渐被学术界抛弃。从整体上来看,单系性是每个分类阶元都追求的基本原则,但接下来我们会看到,不同的具体分类阶元反映出不同层次的生物共性或特性。
02
林奈双名法以及属的意义
当你第一次吃到黄瓤西瓜的时候,你一定会好奇,为什么世上还能有这样的西瓜?有务农经验的长辈这时候也许会解释到“它跟你经常吃到的西瓜不是同一个品种”。换一种更科学的表述方式,我们可以说这种西瓜跟大家经常吃到的红瓤西瓜有着系统性的遗传差异。对于这种不常见的西瓜,我们用“黄瓤西瓜”或“西瓜·黄瓤”来表述就能很容易指出它的与众不同之处来加以区分。
类似的情况在演化研究中再常见不过,例如非洲和亚洲都有体型巨大的大象,但二者又在体型、耳朵大小、象牙长度、趾头数量等方面存在系统性差异。因此,伟大的古生物学家乔治·居维叶(Georges Cuvier,1769-1832年)首次提出将现存的大象分为非洲象和亚洲象两个物种,后来的研究又进一步将非洲象分为非洲草原象和非洲森林象。我们从这里可以看出,演化与生态学家在研究物种多样性和生态功能的时候,绝大多数情况下并不关心两个地理隔离的种群是否形成严格意义上的生殖隔离,而是只要发现了一系列有遗传与生态层面研究意义的,稳定的种群间表型差异,这时候就有必要从相对广泛的生物模式中定义出更具体的物种(Species)。也就是说,在基本认知上,我们可以很容易区分大象这一类生物,但系统性研究要求我们区分出更丰富的遗传演化差异,于是就诞生了林奈双名法。
在林奈之前,物种划分是十分混乱多样的,每个植物学家或动物学家可能都有一套自己的划分方式和命名标准。尤其是为了能全面细致地描述一个物种的特征,当时很多人采用多段词汇堆叠的方式来给物种命名,即多名法(polynomial nomenclature)。随着人类定义出的物种越来越多,这一度带来巨大的混乱。此时林奈引入了一个关键的新思想,即名称的作用只是给一个物种一个独特的标签罢了,大家能知道具体指代的物种是啥就行。于是林奈用两个单词的组合来命名每一个物种,第一个单词代表属名,第二个单词代表种名。只要能保证唯一性,哪怕种和属名都不是精准的特征描述也没问题。例如,鹦鹉科的一只鸟被命名为Psittacula alexandri,意思是“亚历山大的鹦鹉”,以亚历山大大帝的名字命名,亚历山大大帝的军队将东部长尾小鹦鹉引入希腊。
林奈双名法比过去的多名法更容易记忆和使用,并最终取代了它们。这种命名方式历经数百年,直到今天仍能稳定使用下去,这表现了其广泛的科学适用性。
在这套体系里,属(Genus)的作用是至关重要的,因为只要框定了具体属的范围,在此基础上对物种加以区分反倒是一件相对清晰明了的事情。也正因我们无论研究动物植物微生物的时候都普遍使用双名法,属的划分方式必须遵循以下几条十分关键的基本原则:(1)单系性,即一个祖先分类阶元的所有后代被归在一起(即系统发生分析应清楚地证明单系性和作为单独谱系的有效性);(2)合理的紧致性,即属不应该被不必要地扩展;(3)独特性,即符合演化相关的标准或大致共识,即需要从生态学、形态学或生物地理学多方面综合评判。
我们以人类与黑猩猩之间的关系为例,来阐释这一问题。尽管在现今的动物界中,人类和黑猩猩保持着完全相邻的姊妹群体关系,但它们分离的时间已经长达约五百万年。这段时间足以使它们之间形成了稳定的生殖隔离关系,几乎没有任何基因交流。因此,数百万年以来,它们在形态、行为和习性等诸多具体特征上逐渐产生了显著可区分的差异。将它们强行合并到一个属内,将给演化和生态研究带来许多不必要的困扰。而着眼于人属内部,虽然现生的“人类”只有我们智人(Homo sapiens)一个物种,但是尼安德特人(Homo neanderthalensis)等历史上存在过的其他人属物种都曾与智人祖先产生过复杂的基因交流并深刻影响了现代智人的遗传特征,它们的生殖隔离并不严格。而大量的基因组测序分析则向人们揭示了属内物种杂交事件的广泛存在,甚至直接决定了很多新物种的形成。
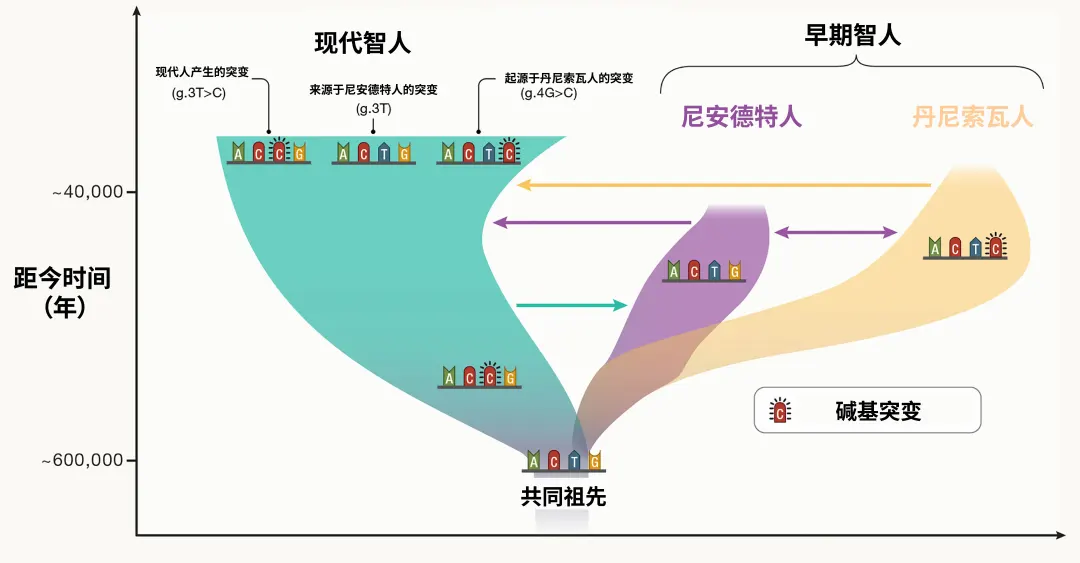
现代智人与早期智人的基因交流。
(图源:https://doi.org/10.1016/j.cell.2023.12.029)
总之,“属”这一概念虽然跟“种”一样做不到尽善尽美的定义,对演化生物学研究来说是十分关键的,它往往反映的是人们普遍认知上对于物种的基本划分方式,就像我们谈到人、老虎、狮子、大象、犀牛、猕猴等常见概念的时候,其实都在生物分类学上完全对应着一个具体的属。换句话说,普遍意义上的生殖隔离大多介于属和种之间,一个属的物种类群要保持足够长的独立演化时间才能被人们接受。
我们可以看到,“属”是一个比较严肃的概念,一个新的生物属一旦被鉴定出来往往会在此后数百年维持不变(属的名字可能会发生变更),但种的划分却相对更加灵活多变,例如将过去定义的一个物种内两个亚种重新区分为两个独立物种是很常见的事情。从遗传独立性的角度讲,属是一个比物种更重要的概念,它在系统发生树中更能代表那些几乎互不交叉的基本末端节点。也就是说,自属往上的系统发生关系,整体上能基本保障树状结构,而从属到种的系统发生关系则往往表现出有内部交叉的网状结构。
03
纲、目、科的生物学基础
从属往上的分类阶元依次是科(Family)、目(Order),以及纲(Class)。分类学家在描述或识别一个科、目或纲的阶元时没有硬性的规则需要遵循。也就是说,这些等级的划分方式在历史上经常发生频繁的变动。例如“纲”的概念在过去曾承担着描述最广泛的共性分类特征的功能,但该功能被德国生物学家恩斯特·海克尔(Ernst Haeckel,1834年-1919年)提出的“门(Phylum)”的概念所取代。
因此我们今天看到的科、目、纲主要是基于分类学家的历史性努力而逐渐固定下来的,所以对于不同生物门类而言,一个“科”所涵盖的遗传多样性和演化时间往往截然不同且毫无可比性。例如,隐翅虫科(Staphylindae)有上千个属,超过63000个现存的已知物种;而人科(Hominidae)只有4个属,8个现生物种。而且人们在纲、目、科之间往往插入诸如下纲、小纲、总目、下目、亚目等数十个相对不常见的阶元。而这些不常见的阶元之间也经常存在变更的情况,如前文提到的平胸总目到古颚下纲的变化过程。
虽然具体的划分方式五花八门,但逐步分类的过程带来了人们对于生命演化逻辑的理解。最基础的就是前文提到的继承自单系共同祖先的同源性特征。而不同的同源特征的产生顺序就反映了演化的基本逻辑。例如,老鹰所属的鹰形目(Accipitriformes)与猫头鹰所属的鸮形目(Strigiformes),在过去的认知中鹰形目是所有陆鸟类的基部类群,而鸮形目则是晚近出现的独立类群,因而老鹰与猫头鹰所具有的肉食性被认为是陆鸟类祖先特征,但依据最新鸟类生命之树,老鹰与猫头鹰应当属于近源物种,共同所属鹰形总目(Accipitrimorphae)鸟类。这样的重新划分使得对于老鹰和猫头鹰的肉食性这一食性的起源的认知可以重新被改写为仅在鹰形总目鸟类的最近共同祖先里才出现的,陆鸟里其他肉食鸟类可能是独立出现了食肉习性。
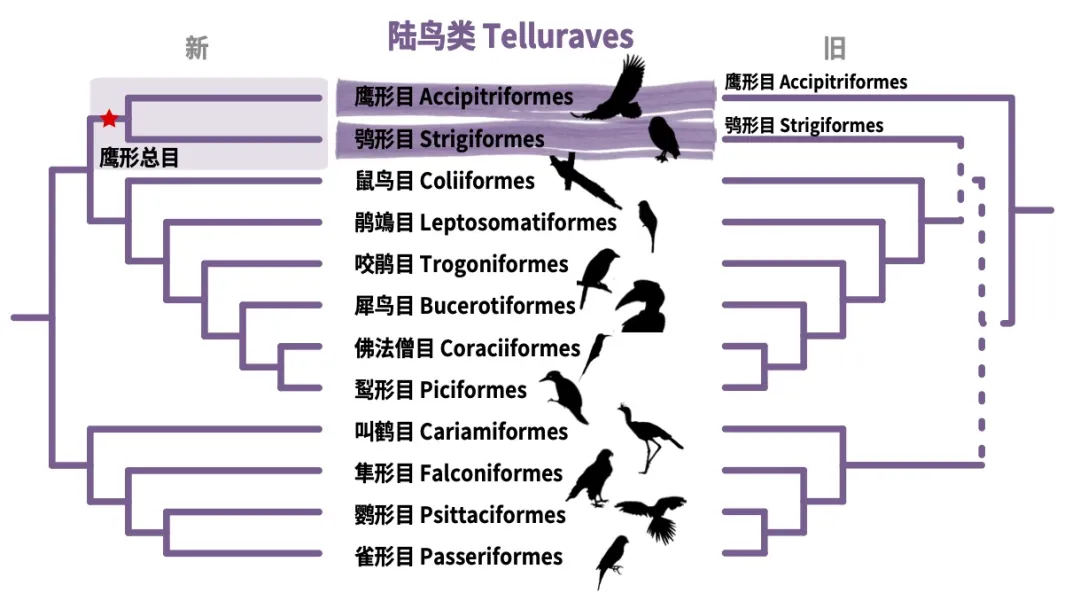
鹰形目与鸮形目亲缘关系的重新划分示意图。左图为新的系统发生关系,支持鹰形目与鸮形目共同组成鹰形总目,右图为以往的系统发生关系,虚线表示新旧系统发生关系的之间的变化。(Josefin Stiller 和 陈光霁绘)
同源特征也可能因退化而消失,例如蛇类等少数爬行动物分支不具备鸟类与爬行动物普遍拥有的四肢结构,但来自古生物化石和胚胎发育的证据都能证明它们是在后续演化历程中丢失了祖先曾有过的四肢发育能力。但我们必须要找到丢失同源性的明确证据才能把不具备更大类群共性特征的某个小类群(蛇下目)放到大类群(爬行纲)的内部。
总的来说,我们在纲、目、科及其内部插入大量其他分类阶元的目的是理解系统发生的真实过程,而是否意味着存在某些生理或者性状特征使得这一类群物种具有适应性优势或巨大的演化潜力(evolvability),或者足以使得其区别于其他类群,这仍有待商榷。例如上述的蛇下目,均具有相同的四肢退化特征。而在传统偶蹄目(Artiodactyla)中,也因新的证据证明了不具备偶蹄特性的鲸豚类与偶蹄目中河马为近源物种,“偶蹄”这一生理特性所代表的偶蹄动物被认为是无效的并系群分类,因此引发了新的分类定义:鲸偶蹄目(Cetartiodactyla)。在这些中间尺度的分类阶元内,各种多变的特征交叉地编码出诸多形态丰富的具体物种,跨越了漫长时空范畴并构建出复杂多样的演化图景。
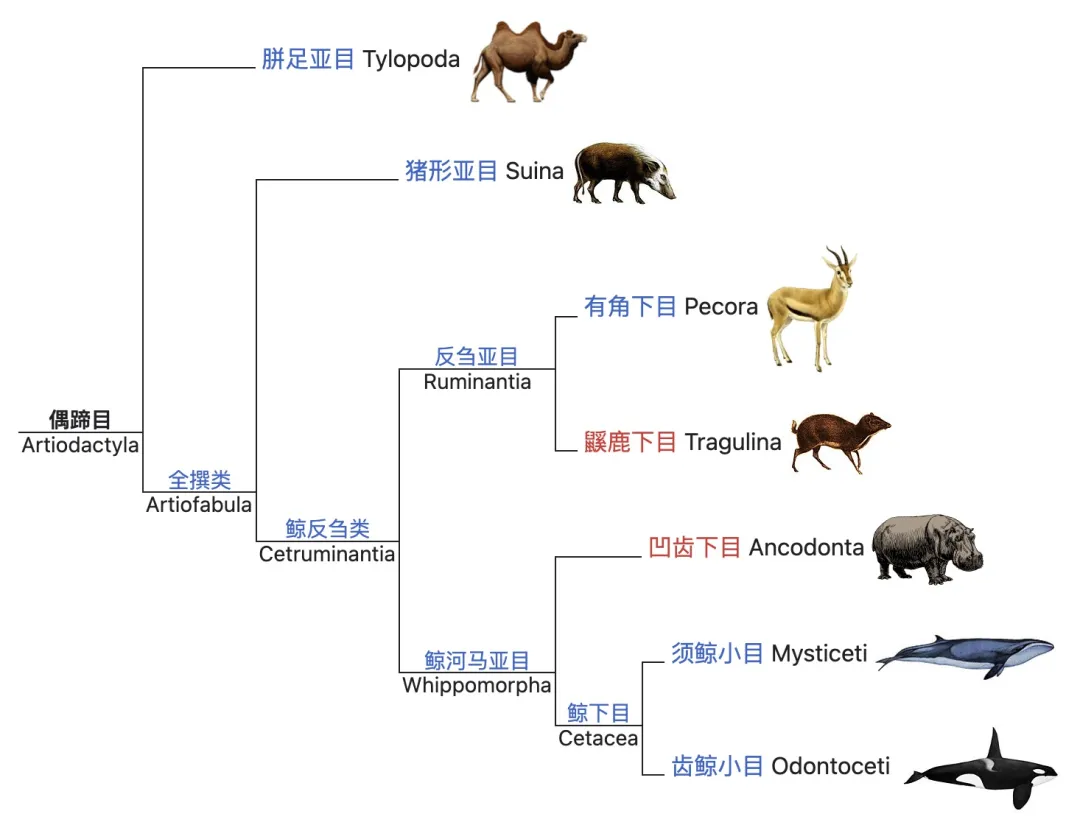
鲸豚类物种属于传统偶蹄目,但因其四足退化而并不具备偶蹄特征。
(图源:维基百科)
04
门与body plan
在很长一段时间里,界(Kingdom)曾经是生物科学分类法中最高的类别。而随着人们对于细胞分子机制的更深入研究,又在界之上添加了作为最高阶元的域。所有细胞生物被划分为了三个域:古菌域、细菌域、真核域。三域划分也存在一定的争议,可以阅读我们早先的内容《当代生物哲学研讨会(四)探索真核生命的起源之迷:一场演化生物学的壮丽征程》。除此之外,域下面界的划分方式也一直处于争论状态,有“三域五界”或“三域六界”之争。每个界代表着截然不同的基本生命形态,不同界的生物甚至不共享完全一致的密码子元件。到目前为止,动物界和植物界的框架是一直稳定的,只是各种单细胞生物的划分归属问题仍处于前沿研究状态,在此不做过多展开。

古生物学鼻祖乔治·居维叶(Georges Cuvier,1769年-1832年)
(图源:维基百科)
界的下一个分类阶元是门(Phylum)。“门”的这一概念的思想构建最早可以追溯到古生物学鼻祖乔治·居维叶(Georges Cuvier,1769年-1832年)。当居维叶试图对动物界的不同种类生物分门别类时发现,有些动物之间似乎无论如何也找不到具体的共享特征,例如将脊椎动物的任何解剖结构都无法跟昆虫有直接对应关系。于是,当居维叶编写《动物界》一书时,将动物分成了四个分支,即脊椎动物、软体动物、有绞动物(节肢动物和环节动物)和植物型动物(居维叶称作“放射虫”,包含棘皮动物、刺胞动物和其它形式),并且指出这些动物都是独立起源演化出来的,没有任何关联。在这种思想的引领下,海克尔首次提出“门”的概念,将其定义为“一个真正的、完全独立的整体,是其所有下辖物种的集合,这些物种从一个共同的原始形式逐渐进化而来,例如,所有脊椎动物。”
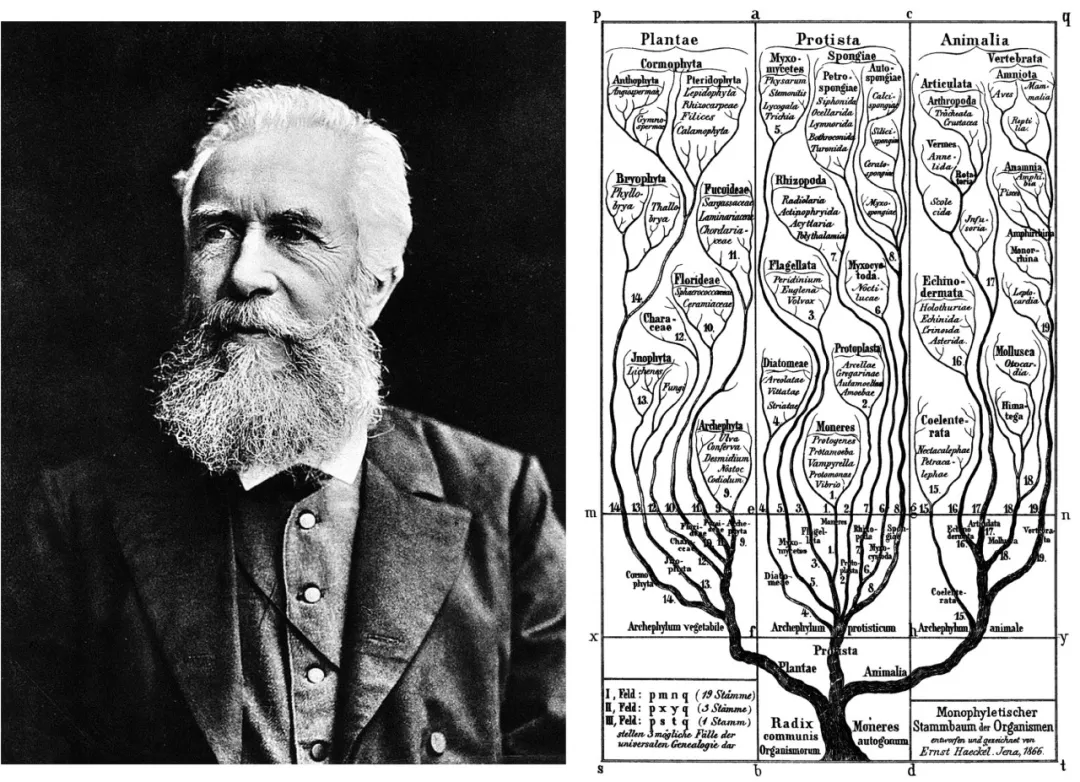
1866年,德国生物学家恩斯特·海克尔对生命进行三域划分
(图源:维基百科)
于是,同一个门的动物共享的基本结构特征组合就被叫做体型呈现(body plan)。定义一个新的门需要强调的是这个门里面的所有生物都无法找到与其他门类生物直接的同源解剖结构。例如,虽然人类和斑马鱼在基本形态上完全不一样,但解剖学会鉴定出一系列同源的骨骼、肌肉、脑区等生理结构,所以只有把它们放在同一个门下面才是合理的。
对于多细胞生物而言,门的划分直接反映的是基础的发育规律。Body plan之所以能在整个门的生物体内都维持保守,是因为它们是产生无数下游特征的结构基础。例如,我们脊索动物门最保守的结构之一就是原肠胚阶段的脊索组织,大多数组织器官的成功分化都需要在脊索的诱导下才能实现,这是发育的因果逻辑决定的。也正因为我们认识到脊椎动物中的脊索结构比脊椎更加基本原始,才将海克尔定义的脊椎动物门变更为脊索动物门,同时将那些无脊椎脊索动物如文昌鱼和海鞘囊括在内。对门类划分的调整都意味着我们对于宏观演化模式有了重大提升,让我们能在更深层次理解演化的原理与内在逻辑。
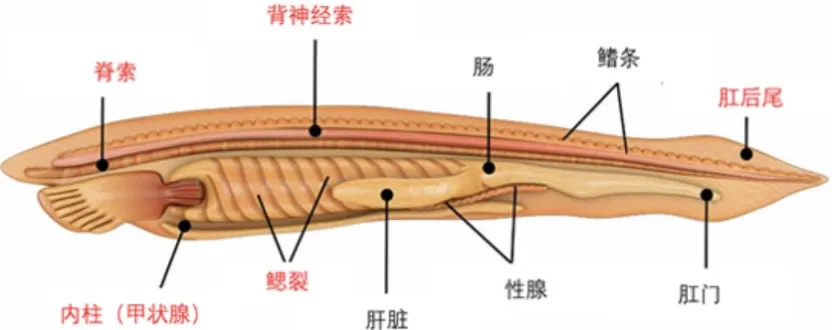
脊索动物门的body plan(标为红色)
(图源:https://biologydictionary.net/vertebrate/)
除了动物界之外,植物界是另一个已被充分研究的领域。几百年来,人们根据维管组织的有无,种子包被方式,花、叶等器官形态划分出了14个现生植物门。
整体上讲,必然是亲缘关系越近的物种大体上长得越像,越能找到更多的类似特征。所以,越是接近科、属级别的划分,越注重具体描述彼此间的形态差异来加以区分;而越接近门、纲级别的划分,则越强调需要用少数保守的共性特征来划分。
也就是说,如果我们发现了一种过去从未见过的生物或化石,哪怕它跟其他所有已知生物相比都看起来长得完全不同,但只要它具备某个门特有的保守特征,我们照样有充分理由把它划分到这个门内。在过去看来,不同门的动植物无法找到直接的同源性证据,但随着发育生物学的深入发展,人们在细胞分化和基因表达调控层面发现不同门生物在解剖结构多样性背后普遍存在深度同源性,这让我们对物种演化有了全新认知。
05
总结
理论上讲,只要得到那棵相对准确的系统发生树(Tree of life),我们完全可以在树的每个二叉节点都命名一个物种类别,这样绝对能保证它们的单系性。当然,真实的物种演化应当并不一定以二叉的形式进行,很有存在多叉或非随机分流等多种复杂状态;所以简单分二叉并进行所有单系群的命名方式必然彻底地丢失了演化的内涵与逻辑。
我们之所以根据脊索等特征定义出脊索动物门,再用脊椎等特征定义出脊椎动物亚门,是因为这样的特征在某个对应的关键祖先节点出现意味着极不寻常的演化事件。在这个过程中,脊椎动物祖先能在某支脊索动物中诞生,是因为脊椎等结构的发育能力需要在脊索组织的诱导下才能构建出来;而脊索动物门属于两侧对称动物则是因为脊索等上游胚胎结构的发育又得建立在两侧对称动物共有的三胚层模式基础上才能实现。因此,我们对于物种门类的划分方式中包含了宏观演化背后的内在逻辑,它不是对于系统发生树进行任意切割都能体现出来的,必须要将树结构和演化趋势进行深度耦合才是有意义的物种分类方式。
总之,界、门、属的概念需要建立在一些生物学普适性原则的基础上提出,而纲、目、科的建立则没有普适性原则。定义一个独立的界需要判断最基本的细胞结构和分子生物学机制的不同,定义一个独立的门需要强调生理形态和解剖结构的不同,定义一个独立的属需要强调对应的生物类群在遗传、演化和生态层面的整体稳定性。
即使存在这些挑战和争议,分类阶元仍然是我们理解和组织生命多样性的重要工具。随着时代的进步,科学技术的不断发展,我们可能会不断审视和调整这些分类阶元的定义与划分。








